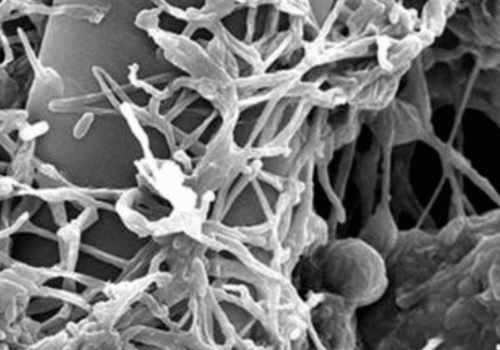
Gliocladium virens: A Mycoparasitic Fungus for Sustainable Disease Management
Overview of the Microbe Gliocladium virens (teleomorph: Trichoderma virens) is a filamentous, soil-dwelling fungus well recognized for its ability to suppress a wide array of...
- 1
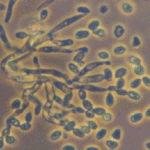
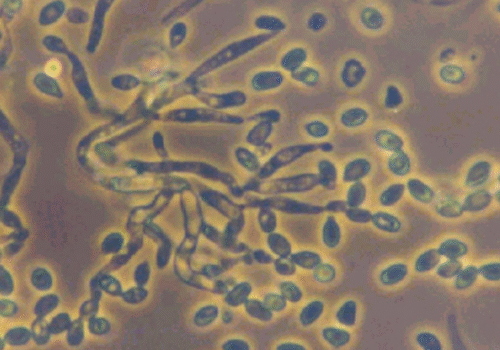
Shows how two fungal strains grow and look under light vs. dark conditions, with measurements of colony size.
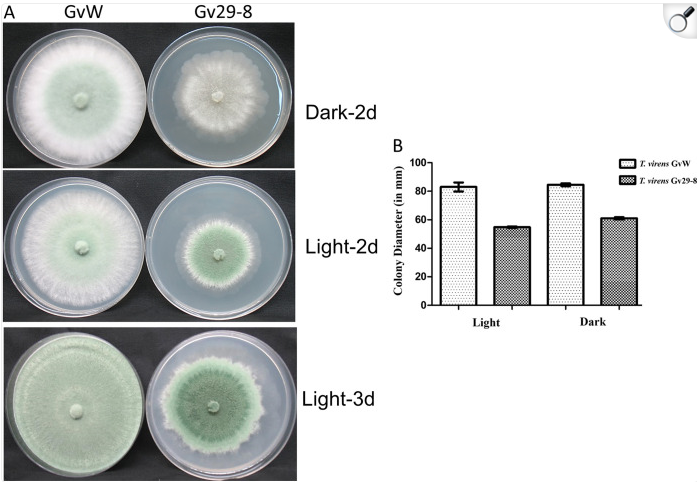
Shows how light and dark affect the growth and appearance of fungal colonies.
- 2
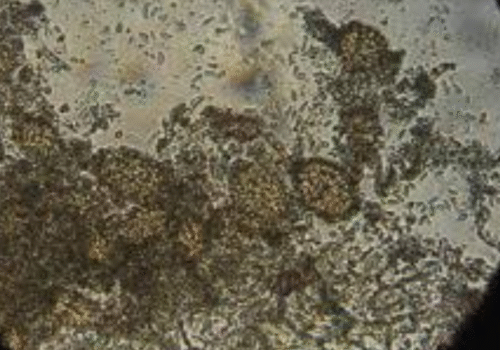
Shows how the fungus S. rolfsii reacts when grown next to two different T. virens strains, comparing their interactions.
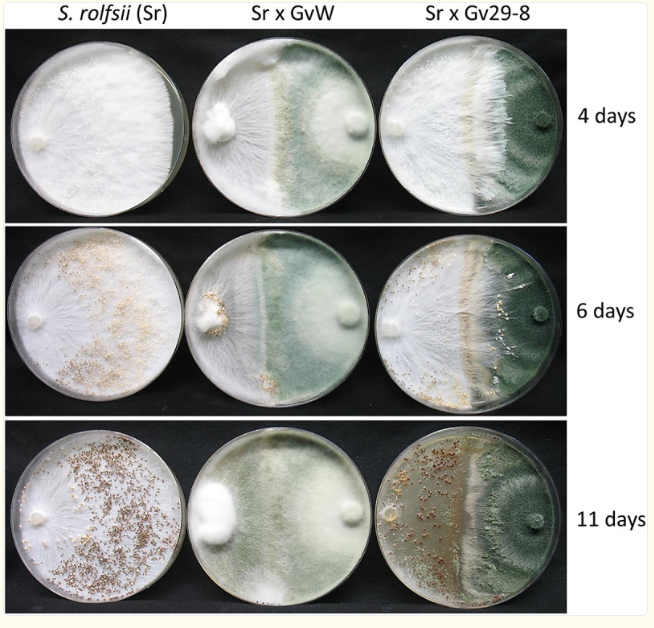
Shows how S. rolfsii interacts with two different T. virens strains when grown side by side.
Overview Table of Gliocladium virens
- Feature
Description
- Scientific Name
Gliocladium virens
- Classification
Ascomycete fungus; Phylum Ascomycota; Order Hypocreales
- Habitat
Rhizosphere, decaying plant material, soil
- Key Functions
Mycoparasitism of phytopathogenic fungi; organic matter decomposition
- Notable Abilities
Secretion of chitinases, glucanases, proteases; spore resilience; rapid colonisation
- Applications
Biocontrol of Rhizoctonia, Pythium, Sclerotium spp.; induced systemic resistance; biofertiliser
- Genetic Engineering Potential
Targeted overexpression of antifungal genes; reporter-strain development for colonisation studies
- Challenges
Variable field efficacy; formulation stability; antagonism by native microflora
- Future Prospects
Metabolome-guided breeding; microbe consortia design; integration with precision agriculture
Overview of the Microbe#
Gliocladium virens (teleomorph: Trichoderma virens) is a filamentous, soil-dwelling fungus well recognized for its ability to suppress a wide array of plant pathogens. As a mycoparasitic and saprophytic organism, G. virens occupies a key ecological niche in the rhizosphere and decomposing organic matter. Formerly classified in the genus Gliocladium, it is now generally accepted under Trichoderma virens based on molecular taxonomy and teleomorphic evidence [1].The fungus is notable for its biocontrol efficacy, environmental safety, and production of secondary metabolites with antibiotic and antifungal activity. Due to these features, G. virens has gained widespread interest in sustainable agriculture, where it serves as an alternative to chemical fungicides for managing soil-borne diseases.

Biocontrol of Soil-Borne Pathogens#
G. virens has demonstrated strong antagonism against a wide range of plant pathogens, including Rhizoctonia solani, Pythium ultimum, Sclerotium rolfsii, and Fusarium oxysporum. Its effectiveness arises from both direct and indirect mechanisms—such as competition for nutrients and space, parasitism of the pathogen, and secretion of antifungal metabolites [2] [3].
Field and greenhouse trials have confirmed its utility in controlling damping-off, seedling blight, and root rots in crops like cotton, tomato, pea, and cucumber [4]. In cotton, for example, formulations containing G. virens have reduced the incidence of seedling diseases caused by Pythium spp. and Rhizoctonia spp. [5].
Commercial products such as SoilGard® are based on G. virens strain G-6, illustrating its practical application in integrated pest management systems..
Mycoparasitism Mechanisms and Lytic Enzymes#
Mycoparasitism is a hallmark trait of G. virens, allowing it to directly attack and penetrate other fungal pathogens. The process involves chemotropic growth toward the host, coiling around the pathogen hyphae, and degrading their cell walls using a suite of lytic enzymes.
The primary enzymes implicated include:
- Chitinases: Break down chitin, a major component of fungal cell walls [6].
- β-1,3-glucanases: Degrade β-glucans in the target fungus [6].
- Proteases: Hydrolyze proteins involved in cell wall integrity [7].
Gene expression studies have shown upregulation of these enzymes during host interaction, confirming their role in pathogenic suppression. Additionally, mutants deficient in chitinase production exhibit reduced biocontrol efficiency, underscoring their importance [6].rmone production works in synergy with its nutrient contributions, amplifying plant growth promotion[1][2].
Induction of Plant Defence Responses#
In addition to attacking pathogens, G. virens enhances the plant’s innate defense mechanisms. It induces systemic resistance, triggering the expression of defense-related genes even in tissues distant from the site of colonization [8].
Studies have demonstrated increased activity of pathogenesis-related (PR) proteins such as peroxidases and glucanases in G. virens-treated plants [9]. This phenomenon, known as Induced Systemic Resistance (ISR), is akin to immunization and offers long-lasting protection against subsequent pathogen attacks.
Moreover, G. virens can modulate hormonal pathways involving jasmonic acid and ethylene, further enhancing the plant’s defense system [10].erefore tends to improve plant vigor and health by multiple routes[2][1].
Secondary Metabolites and Antibiotic Production#
G. virens is a prolific producer of antifungal secondary metabolites, including well-characterized compounds like:
- Gliotoxin: An epipolythiodioxopiperazine with broad-spectrum antifungal and immunosuppressive activity [11].
- Viridin: A furanone with inhibitory effects on fungal spore germination [12].
- Glisoprenins and heptelidic acid: With roles in inhibiting fungal metabolism [13].
These metabolites not only suppress competing fungi but may also participate in signaling and microbial interactions in the rhizosphere. While gliotoxin production is effective, its potential cytotoxicity has raised regulatory concerns regarding certain strains, prompting genome editing approaches to attenuate its expression in commercial biocontrol strains [11]..
Challenges and Future Potential#
Despite its proven potential, deploying G. virens in diverse agricultural settings faces several challenges:
- Strain Variability: Not all strains are equally effective; strain-specific metabolite profiles impact biocontrol consistency [14].
- Environmental Interference: Soil type, temperature, and native microflora can reduce efficacy under field conditions.
- Safety Regulations: The presence of gliotoxin in some strains necessitates thorough biosafety evaluation before large-scale use [11].
Future directions include:
- Genome mining for novel bioactive compounds.
- CRISPR-based editing to enhance beneficial traits.
- Synthetic microbial consortia involving G. virens.· Development of bioformulations with improved shelf life and delivery.
Spotlight on Research: Genome Analysis of Gliocladium virens#
Brief Overview#
A significant study by Lorito et al. [7] titled “Functional genomics of Trichoderma virens reveals pathways for mycoparasitism and secondary metabolite production” provided groundbreaking insights into the genome of G. virens (T. virens).
Key Insights#
Identified ~12,000 protein-coding genes, including those for chitinases, polyketide synthases, and non-ribosomal peptide synthetases.
- Found expanded families of genes linked to oxidative stress tolerance and iron sequestration.
- Annotated gene clusters for gliotoxin and viridin biosynthesis.
- Demonstrated regulatory control via transcription factors such as LaeA and VeA.
Why This Matters#
This genome-scale study offers a molecular blueprint of the fungus’s biocontrol potential and enables precision strain improvement. It also aids in evaluating biosafety concerns through gene knockout models and metabolite regulation strategies.
Summary Table: Spotlight Study#
| Category | Details |
| Lead Researchers | M. Lorito, G.E. Harman, B. Kubicek |
| Affiliations | University of Naples Federico II, Cornell University, TU Wien |
| Research Focus | Genomics, secondary metabolism, mycoparasitism |
| Key Breakthroughs | Identification of 15 secondary metabolite clusters, regulation of ISR pathways |
| Collaborative Efforts | Joint EU-FP7 project with fungal biology consortium |
| Published Work | Current Genetics, 2022 |
| Perspective | Functional genomics to guide safe biocontrol development |
| Publication Date | July 2022 |
| Location | Italy, USA, Austria |
| Key Findings | Genetic map of biocontrol functions and metabolite production |
Conclusion#
Gliocladium virens (syn. Trichoderma virens) exemplifies the shift toward biological and ecological disease management. Its multifaceted mechanisms—mycoparasitism, induction of systemic resistance, and secondary metabolite production—make it a strong contender in the global transition to sustainable agriculture.
However, as with any bioagent, strategic deployment, careful strain selection, and regulatory oversight are critical. The ongoing exploration of its genome and biosynthetic capacity opens new doors for biotechnological innovations, potentially transforming G. virens into a next-generation biofungicide.
References#
- Druzhinina, I. S., et al. (2011). Trichoderma: the genomics of opportunistic success. Nature Reviews Microbiology, 9(10), 749–759. https://doi.org/10.1038/nrmicro2637
- Papavizas, G. C. (1985). Trichoderma and Gliocladium: biology, ecology, and potential for biocontrol. Annual Review of Phytopathology, 23(1), 23–54. https://doi.org/10.1146/annurev.py.23.090185.000323
- Howell, C. R. (2003). Mechanisms employed by Trichoderma species in the biological control of plant diseases: the history and evolution of current concepts. Plant Disease, 87(1), 4–10. https://doi.org/10.1094/PDIS.2003.87.1.4
- Lewis, J. A., & Papavizas, G. C. (1987). Effect of mycoparasite Gliocladium virens on Rhizoctonia solani and the disease it causes in cotton. Phytopathology, 77(5), 843–846. https://doi.org/10.1094/Phyto-77-843
- Lumsden, R. D., Lewis, J. A., & Papavizas, G. C. (1983). Effect of Gliocladium virens on Pythium ultimum, Rhizoctonia solani, and Aphanomyces cochlioides: a comparative study. Phytopathology, 73(6), 773–775. https://doi.org/10.1094/Phyto-73-773
- Harman, G. E., et al. (2004). Chitinolytic and β-1,3-glucanolytic enzymes of Trichoderma virens and their involvement in biocontrol of plant pathogens. Applied and Environmental Microbiology, 70(10), 6083–6091. https://doi.org/10.1128/AEM.70.10.6083-6091.2004
- Lorito, M., et al. (1994). Synergistic interaction between cell wall degrading enzymes and membrane affecting compounds. Phytopathology, 84(10), 950–955. https://doi.org/10.1094/Phyto-84-950
- Yedidia, I., Benhamou, N., & Chet, I. (1999). Induction of defense responses in cucumber plants (Cucumis sativus L.) by the biocontrol agent Trichoderma harzianum. Applied and Environmental Microbiology, 65(3), 1061–1070. https://doi.org/10.1128/AEM.65.3.1061-1070.1999
- Shoresh, M., Yedidia, I., & Chet, I. (2005). Involvement of jasmonic acid/ethylene signaling pathway in the systemic resistance induced in cucumber by Trichoderma asperellum T203. Phytopathology, 95(1), 76–84. https://doi.org/10.1094/PHYTO-95-0076
- Contreras-Cornejo, H. A., et al. (2011). Trichoderma-induced systemic resistance and plant hormesis in maize. Microbial Biotechnology, 4(4), 524–536. https://doi.org/10.1111/j.1751-7915.2010.00247.x
- Gardiner, D. M., et al. (2005). Putative role of gliotoxin in the pathogenicity of Trichoderma virens against Rhizoctonia solani. Phytopathology, 95(9), 1023–1030. https://doi.org/10.1094/PHYTO-95-1023
- Sivasithamparam, K., & Ghisalberti, E. L. (1998). Secondary metabolism in Trichoderma and Gliocladium. In Trichoderma and Gliocladium (Vol. 1, pp. 139–191). Taylor & Francis.
- Mukherjee, P. K., et al. (2013). Secondary metabolites and peptides as key factors in biocontrol. Molecular Plant Pathology, 14(2), 129–138. https://doi.org/10.1111/mpp.12030
- Bailey, B. A., et al. (2008). Diversity and conservation of Trichoderma virens strains used in biocontrol. Mycological Research, 112(8), 928–938. https://doi.org/10.1016/j.mycres.2008.01.024
- Lorito, M., Harman, G. E., & Kubicek, C. P. (2022). Functional genomics of Trichoderma virens reveals pathways for mycoparasitism and secondary metabolite production. Current Genetics, 68(3), 451–465. https://doi.org/10.1007/s00294-022-01233-3