Metarhizium anisopliae: Versatile Entomopathogenic Fungus for Sustainable Pest Control
Overview of the Microbe Metarhizium anisopliae is a globally distributed, filamentous, asexual, soil-dwelling fungus classified under the family Clavicipitaceae, order Hypocreales [1]. First described by...
- Overview of the Microbe
- Entomopathogenic Mechanisms and Pest Management
- Secondary Metabolites and Pharmaceutical Potential
- Plant Bioinoculation and Growth Promotion
- Genetic Diversity and Strain Improvement
- Challenges and Future Potential
- Spotlight on Research: Integrative Taxonomy of the Metarhizium anisopliae Species Complex
- Conclusion
- References
- 1
Evolutionary Tree of Metarhizium Fungi
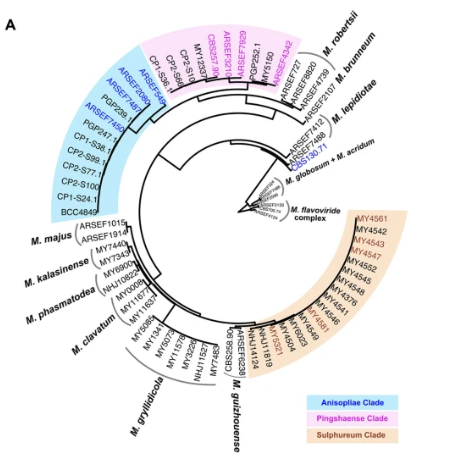
This tree shows how different Metarhizium fungi are related. Fungi with closer branches are more genetically similar.
- 2
Genetic Grouping of Metarhizium Fungi Using Color Blocks
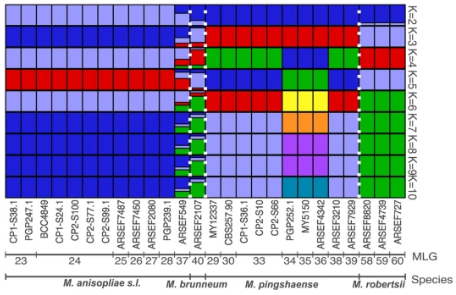
Each colored block represents a different genetic group (K) found in various Metarhizium fungi.
Overview Table of Gluconacetobacter diazotrophicus
- Feature
Description
- Scientific Name
Metarhizium anisopliae (Metch.)
- Classification
Kingdom: Fungi; Phylum: Ascomycota; Class: Sordariomycetes; Order: Hypocreales; Family: Clavicipitaceae
- Habitat
Soil, plant surfaces, insect cadavers
- Key Functions
Entomopathogenesis; production of insecticidal metabolites (destruxins); rhizosphere colonisation
- Notable Abilities
Cuticle penetration via proteases; conidial sporulation; thermotolerance
- Applications
Biopesticides; plant bioinoculants; pharmaceutical metabolite production
- Genetic Engineering Potential
CRISPR-based gene editing; heterologous expression of novel toxins; strain improvement tools
- Challenges
Variable field efficacy; formulation stability; regulatory approval pathways
- Future Prospects
Synthetic biology of destruxin pathways; endophytic strain development; integrated pest management
Overview of the Microbe#
Metarhizium anisopliae is a globally distributed, filamentous, asexual, soil-dwelling fungus classified under the family Clavicipitaceae, order Hypocreales [1]. First described by Metschnikoff in 1879 as Entomophthora anisopliae, it was later reclassified into the genus Metarhizium. Traditionally regarded as the type species of the genus, M. anisopliae infects a broad range of arthropod hosts—over 200 species—causing the characteristic “green muscardine” disease, named after the green conidial coating observed on insect cadavers [2].
The fungus reproduces asexually via conidia that attach to the insect cuticle, germinate, form specialized penetration structures (appressoria), and invade the host. Once inside the hemocoel, the mycelium proliferates and secretes destruxins—cyclic peptides that suppress host immune responses and eventually kill the insect [2]. Its ability to infect diverse insect hosts and persist under varying environmental conditions has made it a cornerstone organism in biological pest control programs worldwide.Historically, many strains were classified under M. anisopliae sensu lato, but recent integrative taxonomic research has resolved this complex into multiple genetically distinct species, including M. robertsii, M. brunneum, and M. majus. According to Kobmoo et al. [3], M. anisopliae sensu stricto corresponds specifically to strain CBS 130.71 and is distinguished by unique morphometric and metabolomic traits. Despite taxonomic reclassification, M. anisopliae s. str. remains a species of interest for its entomopathogenic potential and growing applications in plant–microbe interactions due to its environmental resilience and functional versatility.

Entomopathogenic Mechanisms and Pest Management#
Metarhizium anisopliae infects insects through direct contact, beginning when its asexual spores (conidia) adhere to the host cuticle. The conidia germinate and develop specialized structures known as appressoria, which exert mechanical pressure and secrete cuticle-degrading enzymes—including chitinases, proteases, and lipases—to penetrate the exoskeleton [2]. Once inside the hemocoel, fungal hyphae proliferate and release secondary metabolites such as destruxins, a group of cyclic depsipeptides that suppress insect immunity and contribute to paralysis and death. Under favorable humidity conditions, external sporulation resumes, forming a green layer of conidia on the insect cadaver—giving rise to the term “green muscardine” disease [1].
This entomopathogenic mechanism has enabled M. anisopliae to be deployed successfully against a wide array of insect pests, including termites, thrips, aphids, whiteflies, locusts, agricultural lepidopterans, and even malaria-transmitting mosquitoes [1,2]. A landmark example is the LUBILOSA program, which focused on biological control of locusts and grasshoppers in Africa. While the program ultimately favored Metarhizium acridum for its host specificity, it also reinforced the broader utility and effectiveness of M. anisopliae species in managing diverse pest populations [4].
Commercial formulations of M. anisopliae are available in various forms, including oil-based emulsions, wettable powders, and granules. These are compatible with integrated pest management (IPM) strategies due to their favorable safety profile—being non-toxic to vertebrates and environmentally benign. However, the field efficacy of M. anisopliae can be influenced by factors such as UV exposure, humidity, temperature, and the specific formulation used. Therefore, selecting the appropriate strain and delivery method is essential to optimizing its biocontrol potential.
Secondary Metabolites and Pharmaceutical Potential#
Metarhizium anisopliae produces a wide array of secondary metabolites that contribute to its entomopathogenicity and broader biotechnological potential. Among the most studied are destruxins—cyclic depsipeptides that exhibit potent insecticidal activity by disrupting calcium homeostasis and immune signaling in insect hosts [2]. Destruxins A through E have demonstrated not only paralytic effects on insects but also immunosuppressive and antitumor properties in laboratory models [2, 5].
Yin et al. (2021) showed that destruxin A directly interacts with insect proteins such as SEC23 and TEME214, impairing vesicle transport and immune function, ultimately accelerating host mortality. Importantly, these compounds degrade rapidly post-mortem, minimizing risks of environmental accumulation and toxicity to non-target organisms.
In addition to destruxins, M. anisopliae is known to produce other metabolites including protease inhibitors, swainsonine, and a range of bioactive alkaloids and peptides. These compounds have demonstrated antimicrobial, antifungal, and cytotoxic effects against tumor cell lines, suggesting potential pharmaceutical applications [6].Recent advancements in molecular biology have further expanded the potential of M. anisopliae through genetic engineering. For example, transgenic strains have been developed that express insect-specific neurotoxins or anti-malarial antibodies, significantly increasing virulence while preserving host specificity [5]. These innovations open new avenues not only for enhanced pest control but also for the potential therapeutic exploitation of fungal metabolites in medicine and biotechnology.
Plant Bioinoculation and Growth Promotion#
In addition to its well-established role as an entomopathogen, Metarhizium anisopliae functions as a plant-beneficial endophyte capable of colonizing both root and aerial tissues. Multiple studies have documented its successful endophytic association with crops such as maize, rice, soybean, cotton, and tomato, where it enhances plant development through improved nutrient uptake and increased tolerance to abiotic stressors like drought and salinity [7, 8].
Mechanistically, M. anisopliae supports plant growth by producing phytohormones—including indole-3-acetic acid (IAA) and gibberellins—and by modulating plant defense-related gene expression to induce systemic resistance (ISR) [8]. It also contributes to better mineral uptake, particularly nitrogen and phosphorus, due to its ability to alter root architecture and exude plant-growth-promoting compounds.
In a hydroponic maize experiment, Nowak et al. [9] demonstrated that M. anisopliae colonized root systems systemically and led to substantial increases in plant height, root length, chlorophyll content, and total biomass—exceeding control values by over 60%. These effects were consistent across different environmental conditions, highlighting the fungus’s adaptability and utility in sustainable agriculture systems.
The dual role of M. anisopliae as a biological control agent and a plant bioinoculant makes it a highly promising tool in integrated agroecological practices. Its multifunctionality reduces reliance on synthetic fertilizers and pesticides, offering a microbe-centered solution to enhance crop productivity while maintaining environmental integrity.
Genetic Diversity and Strain Improvement#
Taxonomically, M. anisopliae was long treated as a catch-all species. However, genomic advances have revealed significant cryptic diversity. Kobmoo et al. [3] applied integrative taxonomy—combining phylogenomics, morphometrics, metabolomics, and virulence data—to delineate the M. anisopliae species complex.
Key findings included:
- M. anisopliae sensu stricto corresponds to strain CBS 130.71, previously misidentified.
- Discovery of three new species: M. neoanisopliae, M. hybridum, and M. parapingshaense.
- Each species exhibits unique metabolomic profiles and conidial morphometrics.
- Field-deployed strains represent stable clonal populations adapted to specific ecological niches.
This diversity underscores the importance of strain verification in biocontrol programs and opens avenues for genetic improvement. With tools like CRISPR and synthetic biology, enhancements in spore viability, virulence, and metabolite production are increasingly feasible.ongisporum fits well into IPM frameworks for sustainable and ecologically friendly pest control.
Challenges and Future Potential#
Challenges#
- Taxonomic confusion: Misidentification affects efficacy and regulation.
- Environmental limitations: Sensitivity to UV and humidity can reduce performance.
- Formulation issues: Conidia degrade in field conditions; oil-based or encapsulated formulations are needed.
- Non-target effects: Though generally safe, effects on non-target insects (e.g., pollinators) must be evaluated.
Future Directions#
- Dual-purpose bioinoculants: Integrating pest control and plant growth enhancement.
- Genomic-guided improvement: Targeting specific traits like spore adhesion and stress tolerance.
- Bioremediation: Emerging evidence shows some Metarhizium species can degrade heavy metals and hydrocarbons.
- Climate-smart agriculture: Especially relevant in arid and saline-prone regions due to the fungus’s stress tolerance.
Spotlight on Research: Integrative Taxonomy of the Metarhizium anisopliae Species Complex#
Brief Overview#
Kobmoo et al. [3] conducted the most comprehensive taxonomic revision of the M. anisopliae complex to date. Their study used:
- Phylogenomics (whole-genome sequencing)
- Morphometrics (conidia size, shape)
- Metabolomics (LC-MS profiles)
- Virulence assays (against Galleria mellonella)
This multi-pronged approach redefined species boundaries and led to the discovery of three novel species.
Key Insights#
- M. anisopliae sensu stricto aligns with CBS 130.71 and is genetically distinct from many commercial strains.
- The new species display distinct conidial dimensions and secondary metabolite profiles.
- Field isolates show low genetic variability, indicating strong clonal persistence.
Why This Matters#
This research allows:
- Greater regulatory clarity for microbial biopesticides
- More accurate strain identification for biocontrol
- Tailored formulations based on species-specific traits
Summary Table: Spotlight Study#
| Category | Details |
| Lead Researchers | Noppol Kobmoo et al. |
| Affiliations | BIOTEC, Thailand and international collaborators |
| Research Focus | Taxonomic clarity of M. anisopliae complex |
| Key Breakthroughs | Identification of three new species; redefinition of s. str. |
| Published Work | IMA Fungus, 15(1): 30 |
| Publication Date | 2024 |
| Location | Global strain coverage; coordinated from Thailand |
| Key Findings | Genomic and phenotypic resolution for effective biocontrol |
Conclusion#
Metarhizium anisopliae exemplifies nature’s multifunctionality—serving both as an insect pathogen and a plant symbiont. Its complex infection mechanisms, biostimulatory potential, and evolving genomic insights offer exciting prospects for sustainable agriculture, environmental restoration, and possibly pharmaceutical innovation. However, unlocking its full potential hinges on precise taxonomy, strain-specific application, robust formulation, and regulatory harmonization. As agriculture and environmental health intersect, M. anisopliae is poised to lead a microbial revolution in eco-friendly innovation.
References#
- Skrobek, A., Shah, F. A., & Butt, T. M. (2008). Destruxin production by the entomogenous fungus Metarhizium anisopliae in insects and factors influencing their degradation. Biocontrol Science and Technology, 18(8), 781–793. https://doi.org/10.1007/s10526-007-9077-1
- Yin, F., Xiao, M., Berestetskiy, A., & Hu, Q. (2021). The Metarhizium anisopliae toxin, destruxin A, interacts with the SEC23A and TEME214 proteins of Bombyx mori. Journal of Fungi, 7(6), 460. https://doi.org/10.3390/jof7060460
- Kobmoo, N., Mongkolsamrit, S., Khonsanit, A., Cedeño-Sanchez, M., Arnamnart, N., Noisripoom, W., … & Luangsa-ard, J. J. (2024). Integrative taxonomy of the Metarhizium anisopliae species complex, based on phylogenomics combined with morphometrics, metabolomics, and virulence data. IMA Fungus, 15(1), 30. https://doi.org/10.1186/s43008-024-00154-9
- Lomer, C. J., Bateman, R. P., Johnson, D. L., Langewald, J., & Thomas, M. (2001). Biological control of locusts and grasshoppers. Annual Review of Entomology, 46, 667–702. https://doi.org/10.1146/annurev.ento.46.1.667
- St. Leger, R. J., Wang, C., & Fang, W. (2016). New perspectives on insect pathogens. Fungal Biology Reviews, 30(3), 67–72. https://doi.org/10.1016/j.fbr.2016.06.001
- Wu, C. C., Chen, T. H., Liu, B. L., Wu, L. C., Chen, Y. C., Tzeng, Y. M., & Hsu, S. L. (2013). Destruxin B Isolated from Entomopathogenic Fungus Metarhizium anisopliae Induces Apoptosis via a Bcl‐2 Family‐Dependent Mitochondrial Pathway in Human Nonsmall Cell Lung Cancer Cells. Evidence‐Based Complementary and Alternative Medicine, 2013(1), 548929. https://doi.org/10.1155/2013/548929
- Liu, Y., Yang, Y., & Wang, B. (2022). Entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae play roles of maize (Zea mays) growth promoter. Scientific reports, 12(1), 15706. https://doi.org/10.1038/s41598-022-19899-7
- Chaudhary, P. J., BL, R., Patel, H. K., Mehta, P. V., Patel, N. B., Sonth, B., … & Kaushik, P. (2023). Plant growth-promoting potential of entomopathogenic fungus Metarhizium pinghaense AAUBC-M26 under elevated salt stress in tomato. Agronomy, 13(6), 1577. https://doi.org/10.3390/agronomy13061577
- Nowak, M., Bernat, P., & Różalska, S. (2024). Mutual interaction of the entomopathogenic and endophytic fungus Metarhizium anisopliae with zearalenone as a native component of crude Fusarium extract. Scientific Reports, 14, Article 22493. https://doi.org/10.1038/s41598-024-73022-6