Bacillus thuringiensis as a Green Microbe: Advances, Challenges, and Future Prospects for Sustainable Agriculture
Abstract Bacillus thuringiensis (Bt) has emerged as a cornerstone of sustainable pest management, celebrated as a “green” microbial insecticide in agriculture. This review provides a...
- Abstract
- Introduction
- Historical Overview of Bacillus thuringiensis
- Molecular Biology of Bt and Mode of Action of Cry and Cyt Proteins
- Bt in Sustainable Agriculture: Biopesticides and Transgenic Crops
- Ecological and Environmental Safety Profile of Bt
- Emerging Challenges: Insect Resistance and Regulatory Concerns
- Innovations and Future Prospects in Bt Biotechnology
- Conclusion
- References
- 1
Powdery Mildew Infection on Melon Leaves and Fungal Structures

(A) Melon leaf showing white, powdery mildew patches caused by fungal growth. (B) Microscopic view of spore chains on leaf surface.
- 2
Fungal Fruiting Bodies and Spore Structures on Infected Plant Tissue
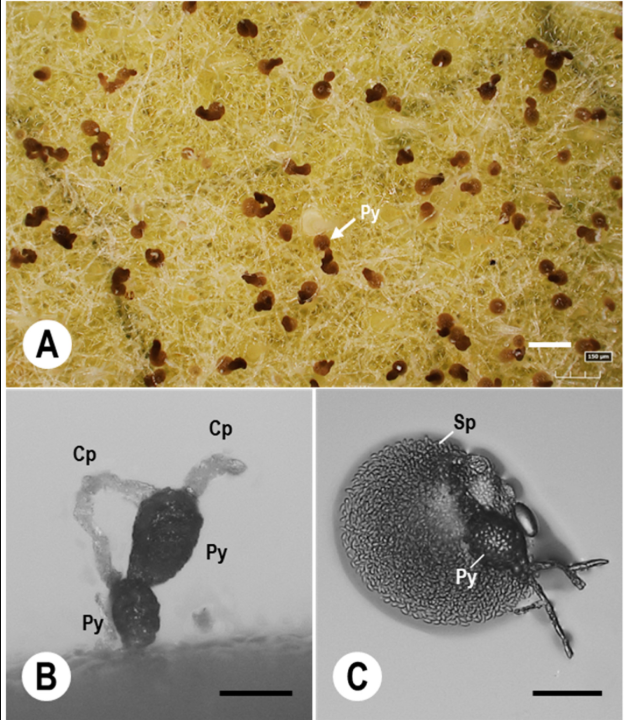
(A) Numerous dark, round fungal fruiting bodies (pycnidia) scattered across infected plant surface. (B) Close-up showing pycnidia (Py) with connecting cells (Cp). (C) Detailed view of a spore-filled pycnidium (Py) with spores (Sp) visible inside.
Overview Table of GAmpelomyces quisqualis
- Feature
Description
- Scientific Name
Ampelomyces quisqualis (Ces.) Rabenh. (1851)
- Classification
Phylum: Ascomycota; Order: Pleosporales; Family: Phaeosphaeriaceae
- Habitat
Broadly distributed as a hyperparasite on powdery mildew fungi across diverse crops and climates
- Key Functions
Mycoparasitism of Erysiphales (powdery mildew); suppression of disease spread
- Notable Abilities
Production of cell-wall–degrading enzymes; sporulation within host structures; high host specificity
- Applications
Biocontrol of powdery mildews in greenhouse and field crops; organic‐approved biofungicide formulations
- Genetic Engineering Potential
Targets: genes for chitinase and β-1,3-glucanase; tools: CRISPR/Cas9 to enhance enzyme expression
- Challenges
Variable field efficacy; humidity dependence; production scale‐up; regulatory registration
- Future Prospects
Strain improvement via synthetic biology; AI‐driven fermentation optimisation; integration in circular agrifood systems
Abstract#
Bacillus thuringiensis (Bt) has emerged as a cornerstone of sustainable pest management, celebrated as a “green” microbial insecticide in agriculture. This review provides a comprehensive overview of Bt, from its historical discovery over a century ago to its modern applications and future potential. We recount the early discovery of Bt and its development as a biopesticide, including key milestones such as the identification of insecticidal crystal (Cry) proteins. We then examine Bt’s molecular biology and mode of action in detail, highlighting the diverse Cry and Cyt toxins and their specific activities against insect pests. The deployment of Bt in sustainable agriculture is discussed in depth—both as formulated biopesticides and through genetically modified (GM) Bt crops. Evidence of the ecological safety of Bt is reviewed, noting its high specificity and generally minimal non-target impacts, alongside considerations of environmental fate. We address emerging challenges, particularly the evolution of insect resistance to Bt toxins and the regulatory and public perception issues surrounding GM Bt crops. Innovations and future prospects are also explored, including novel toxin genes, biotechnological improvements to enhance efficacy and durability, and expanded uses of Bt (such as plant growth promotion and bioremediation). Throughout, we underscore that Bacillus thuringiensis exemplifies a successful synergy between microbiology and agriculture—advancing crop protection in an environmentally sustainable manner.
Keywords: Bacillus thuringiensis; biopesticides; Cry protein; Bt crops; sustainable agriculture; insect resistance; ecological safety

Introduction#
Sustainable agriculture seeks to maintain high crop productivity while minimizing harm to the environment and human health[1][2]. A major challenge in agriculture has been the management of insect pests in an ecologically sound way. For decades, pest control relied heavily on synthetic chemical insecticides, which, while effective, often cause collateral damage to beneficial organisms, foster pest resistance, and leave harmful residues[3][4]. In this context, Bacillus thuringiensis (Bt), a naturally occurring soil bacterium with insecticidal properties, has gained prominence as a green alternative. Bt produces protein toxins that specifically target certain insects while being largely innocuous to non-target organisms and humans[5][6]. These characteristics have made Bt a linchpin of biological pest control and a key component of integrated pest management (IPM) strategies.
This review provides a detailed examination of Bacillus thuringiensis and its role in sustainable agriculture. We begin with a historical overview of Bt’s discovery and early uses as a microbial insecticide. We then delve into the molecular biology of Bt, describing its toxin families (Cry, Cyt, and others) and their mode of action against insect pests. The use of Bt in sustainable agriculture is discussed in two main contexts: (1) Bt as a biopesticide applied in sprays or formulations, and (2) Bt as expressed in genetically modified crops (Bt crops). We review evidence on the environmental safety of Bt, given its host specificity, and discuss any ecological impacts. Emerging challenges, notably insect resistance to Bt and regulatory/public acceptance issues, are analyzed. Finally, we explore innovations and future prospects in Bt-related biotechnology, including new toxin development, improved deployment methods, and novel applications beyond insect control. By consolidating recent research (primarily from the last 5–10 years, along with foundational literature), this article aims to provide a comprehensive, up-to-date resource on Bacillus thuringiensis as a sustainable tool for agriculture.
Historical Overview of Bacillus thuringiensis#
The story of Bacillus thuringiensis began at the turn of the 20th century. In 1901, Japanese biologist Shigetane Ishiwatari isolated a bacterium as the cause of a disease (“sotto”) that was killing silkworm larvae; he named it Bacillus sotto, not yet realizing it was a new species[7]. A decade later in 1911, German microbiologist Ernst Berliner independently rediscovered the organism while investigating a fatal disease of flour moth caterpillars (Ephestia kuehniella) in the Thuringia region of Germany[7][8]. Berliner named it Bacillus thuringiensis after Thuringia, and it was initially regarded as a threat to silkworm cultivation before its potential as an insecticide was recognized[9]. Early on, scientists observed that the bacterium’s pathogenicity to insects was due to a toxic factor, presaging the discovery of Bt’s crystal toxins[10][11].
By the 1920s and 1930s, B. thuringiensis was being considered for practical pest control. In 1927, Berliner’s Bt strain was tested in field trials against the European corn borer, demonstrating promise as a biocontrol agent[11]. The first commercial Bt product, named Sporeine, was produced in France in 1938[12]. Sporeine was a sprayable formulation of Bt spores and crystalline toxins, used to combat lepidopteran pests. However, broader adoption of Bt sprays did not take off until the mid-20th century. In the 1950s, interest in microbial insecticides resurged: Eastern European countries and the United States developed Bt-based products (e.g. trade names like Thuricide appeared in the U.S.), bringing Bt to market as a registered insecticide[13][14].
Scientific breakthroughs in the mid-20th century solidified understanding of how Bt works. In 1953, researcher Christopher Hannay observed under the microscope that Bt during sporulation produces parasporal crystal inclusions[11]. Soon after, Angus (1956) experimentally demonstrated that these crystalline inclusions were the source of Bt’s insecticidal action[15]. The crystal was found to be proteinaceous and toxic when ingested by susceptible insects. Pioneering genetic studies followed in the 1970s: Zakharyan and co-workers (1977) provided evidence that the genes encoding the crystal toxins reside on plasmids in Bt[16]. González et al. (1982) confirmed this by curing Bt of its plasmids, which eliminated toxin production, proving that toxin genes were plasmid-borne[16]. The biotechnology era propelled Bt research to new heights. In 1981, Schnepf and Whiteley cloned and expressed the first Bt toxin gene (a cry gene from B. thuringiensis kurstaki HD-1) in Escherichia coli, demonstrating that the gene product was toxic to insect larvae[17]. This landmark achievement opened the door to engineering plants with Bt genes. By the mid-1990s, the first transgenic Bt crops became a reality: Bt potato was commercialized in 1995 (effective against Colorado potato beetle), followed by Bt cotton and Bt corn in 1996[18]. Bt cotton, in particular, reached farmers’ fields in 1996 and rapidly gained approval and use in multiple countries[18]. Thus, within less than a century of its discovery, Bacillus thuringiensis evolved from an obscure insect pathogen into a globally important tool for crop protection.
Molecular Biology of Bt and Mode of Action of Cry and Cyt Proteins#
Bt Toxins: Diversity and Classification#
The insecticidal power of B. thuringiensis resides in the array of toxins it produces. Chief among these are the Cry (crystal) proteins and Cyt (cytolytic) proteins, collectively known as δ-endotoxins. These proteins are typically produced during sporulation, forming parasporal crystalline inclusions within the bacterium’s sporangium[19]. Hundreds of distinct Bt toxin genes have been identified, reflecting a remarkable diversity. As of 2020, over 800 cry genes had been described, classified into at least 75 families (Cry1, Cry2, … Cry75), along with ~40 cyt genes grouped into 3 families[20]. Bt also produces Vip (vegetative insecticidal proteins) and Sip (secreted insecticidal proteins) during its vegetative growth phase, which expand its pesticidal repertoire[21].
Historically, Cry toxins were first grouped by their spectrum of activity. The classic classification by Höfte and Whiteley (1989) defined four main Cry classes based on amino acid sequence homology and target insect orders[22]. Cry1 toxins are active primarily against Lepidoptera (caterpillars of butterflies and moths), Cry2 toxins affect Lepidoptera and Diptera (flies, mosquitoes), Cry3 toxins target Coleoptera (beetles), and Cry4 toxins kill Diptera (especially mosquito larvae)[22]. A fifth class “Cry5” (sometimes called CryV in older literature) was later designated for toxins active against nematodes (roundworms)[23]. This structure is somewhat simplified, as continued discovery has greatly expanded known toxin types. In modern nomenclature, each novel Cry protein is assigned a number-letter combination (e.g. Cry1Ac, Cry2Ab) based on hierarchical amino acid similarity criteria[24]. Cyt toxins, by contrast, are a separate class of β-sheet pore-forming toxins; Cyt proteins (e.g. Cyt1A, Cyt2B) are known for their role in mosquito-active Bt strains and can synergize with Cry toxins to enhance potency against dipteran larvae[25][26]. The vegetative toxins include Vip3A (a single-chain protein toxic to lepidopterans) and the binary Vip1/Vip2 which jointly affect certain coleopteran and hemipteran insects[27]. Table 1 summarizes the major classes of Bt toxins and their typical insect targets.
Table 1. Classes of B. thuringiensis Insecticidal Toxins and Their Target Insect Groups
| Bt Toxin Class | Representative Toxins (Examples) | Primary Target Insect Orders |
| Cry1 family | Cry1A (e.g. Cry1Ac, Cry1Ab) | Lepidoptera – caterpillars (butterflies, moths)[28]. Notable target pests: Helicoverpa spp., Ostrinia nubilalis (European corn borer). |
| Cry2 family | Cry2A (e.g. Cry2Ab) | Lepidoptera and Diptera – caterpillars and fly/mosquito larvae[28]. (Dual activity on moths and mosquitoes). |
| Cry3 family | Cry3A, Cry3Bb | Coleoptera – beetle larvae (e.g. Diabrotica rootworms, Leptinotarsa potato beetle)[28]. |
| Cry4 family | Cry4A, Cry4B, Cry11 | Diptera – mosquito and black fly larvae[28]. Used in B. thuringiensis israelensis for mosquito control. |
| Cry5/Cry6 (Nematocidal) | Cry5B, Cry6A | Nematodes – roundworms (some species)[23]. Experimental use against plant-parasitic or vector nematodes. |
| Cyt family | Cyt1A, Cyt2B | Diptera (mosquitoes, flies) – usually synergistic with Cry toxins[25]. Cyt toxins augment mosquito larvicidal activity and help delay resistance[26]. |
| Vip1/Vip2 (binary toxins) | Vip1Aa + Vip2Ab | Coleoptera and Hemiptera – (binary toxin requires both proteins)[27]. Investigated for beetle pests and sap-sucking insects. |
| Vip3 family | Vip3A (e.g. Vip3Aa) | Lepidoptera – caterpillars of various species[27]. Used in some transgenic crops to broaden lepidopteran control spectrum. |
(Data compiled from sources: Kumar et al. 2021[29][27]; Butko 2003; Höfte & Whiteley 1989.)
In addition to the well-known toxins above, certain B. thuringiensis strains produce Parasporins – these are crystal proteins that are not insecticidal but have cytotoxic effects on cancer cells, underlining the biochemical diversity of Bt toxins[30]. However, parasporins are outside the scope of agricultural use and are being studied for medical applications.
Mode of Action of Bt Toxins#
Bacillus thuringiensis toxins have a unique mode of action that underpins their specificity and potency. The primary pathway of killing is as follows: ingestion, activation, binding, pore formation, and cell lysis[31][32]. When a susceptible insect larva feeds on a Bt-spore/crystal mixture (either on a sprayed leaf or a transgenic plant), the Cry proteins are solubilized in the alkaline pH of the insect midgut. In Lepidoptera, for example, midgut pH can be ~9.5, which dissolves the crystalline endotoxin. Gut proteases then cleave the protoxin into an active toxin core[31]. The activated toxin binds to specific receptors on the brush-border membrane of midgut epithelial cells (receptors may include cadherin-like proteins, aminopeptidases, and other membrane proteins in the insect gut)[33]. Upon binding, several Cry toxin molecules insert into the cell membrane and oligomerize to form pores (ion channels). This pore formation causes osmotic imbalance, cell swelling, and eventual lysis of the midgut epithelial cells[33][32]. In essence, the midgut lining is disrupted—histopathological changes include swelling and bursting of columnar cells, leakage of gut contents, and paralysis of the digestive tract[34][35]. Affected larvae cease feeding within hours and typically die from a combination of midgut tissue necrosis and septicemia (gut bacteria invading the hemocoel) or starve due to gut paralysis[36][37].
The timeline of Bt toxin action in a lepidopteran larva can be rapid. Studies show that within minutes of ingestion, physiological changes occur: increased ion flux in gut cells, loss of membrane potential, and beginning of cell breakdown[38]. Within a few hours, larvae become sluggish (gut paralysis sets in), and within 1–2 days, death ensues[36]. This mode of action is often described as causing a “leaky gut” in the insect. Importantly, the specificity of Bt toxins arises from the requirement of specific gut conditions and receptors—if an organism lacks the appropriate enzyme conditions to activate the toxin or the receptors for toxin binding, the Bt toxin will have no effect[39][40]. For example, Cry1 toxins bind receptors present in caterpillar guts but not in mammals, which is why Bt is selectively toxic to those insects and safe to vertebrates.
The Cyt toxins, produced by certain Bt strains (notably B. thuringiensis israelensis used for mosquito control), have a somewhat different action. Cyt proteins can directly insert into cell membranes and form pores without needing specific receptors, but they are less potent alone. Their main value is synergistic: Cyt1A, for instance, can bind to and sequester membranes in a way that prevents mosquitoes from developing resistance to Cry toxins, essentially delaying resistance by providing an alternative binding mechanism[25][26]. In mosquito larvae, Cyt1A works alongside Cry4 and Cry11 toxins to ensure robust control even if one toxin’s receptor is altered by resistance mutations[26].
Beyond the gut epithelial attack, some Cry toxins can trigger additional pathways like septicemia. After midgut rupture, B. thuringiensis spores can invade the insect’s body cavity and multiply, contributing to insect death. However, modern Bt biopesticide formulations often exclude live spores to focus on the toxin action and address regulatory concerns (spore-free formulations are sometimes preferred)[41]. Nonetheless, the combination of spore and toxin in nature is an effective one-two punch: toxin creates an entry, and the bacterium exploits the opportunity to infect the host.
In summary, the mode of action of Bt’s Cry and Cyt proteins is well-characterized and can be seen as an elegant example of targeted biochemistry: the toxin exploits specific vulnerabilities in the insect gut that are absent in non-target organisms. Figure 1 illustrates the sequence of events in a susceptible insect midgut after Bt toxin ingestion (from activation to cell lysis and gut paralysis). This specificity not only makes Bt effective but also underlies its reputation as an environmentally friendly insecticide.
(Figure 1. Mode of action of Bt Cry toxins in insect midgut – not shown in text.)
Bt in Sustainable Agriculture: Biopesticides and Transgenic Crops#
Bt as a Microbial Biopesticide#
The most traditional application of Bacillus thuringiensis in agriculture is as a biopesticide, wherein formulations of the bacterium (or its spores and toxins) are applied to crops to kill pests. Bt biopesticides have been used since the 1940s on a modest scale, but their use expanded significantly from the 1960s onward as awareness of the drawbacks of chemical pesticides grew. Bt is often called a “microbial insecticide” – farmers spray it much like a chemical insecticide, but its active ingredient is biological (living bacteria and/or their natural toxin product).
Bt biopesticide products typically contain spores and toxin crystals of specific B. thuringiensis strains targeted at particular pest groups. B. thuringiensis kurstaki (Btk) strain HD-1, for instance, is widely used against lepidopteran pests (caterpillars) on vegetables, fruits, and forestry (it controls pests like cabbage worms, loopers, and gypsy moth larvae). B. thuringiensis israelensis (Bti) is specialized for dipteran pests and is employed in mosquito control programs to kill mosquito and blackfly larvae in water bodies[42]. Another strain, B. thuringiensis tenebrionis (also known as Bt var. san diego), is used to manage certain beetles – notably the Colorado potato beetle on potatoes[43]. These products have narrow spectra, which is advantageous for integrated pest management: they target key pests while sparing beneficial insects like pollinators and predators (e.g. Btk will kill caterpillars on a crop but not harm bees or ladybugs)[5].
The commercial success of Bt biopesticides is significant. By the 1990s, Bt-based products constituted the largest share of biopesticide sales worldwide[6]. They remain a mainstay of organic agriculture, since Bt sprays are one of the few tools available to organic farmers for effective pest control (organic standards permit Bt since it is a natural bacterium, not a synthetic chemical). Many dozens of Bt formulations exist globally – for example, trade names like Dipel, Thuricide, VectoBac, XenTari, and Novodor correspond to different Bt strains/formulations for different pest targets. According to one analysis, biopesticides derived from Bacillus species (predominantly Bt) represent the most significant class of commercial biocontrol products[6]. Their popularity stems from several factors: high efficacy on target pests, safety to non-targets and consumers, compatibility with other IPM tactics, and the fact that Bt residues degrade quickly in the environment (the proteins are biodegradable, and the spores persist but are not invasive)[5][44].
Despite these advantages, there are practical considerations and challenges in using Bt biopesticides. One issue is their relative specificity and slower action compared to broad-spectrum chemicals. For instance, a Bt spray might take a day or two to kill caterpillars (as the pests must ingest it), whereas a contact poison could kill within hours. Also, UV sunlight can degrade Bt toxins, and rain can wash spores off leaves, so formulations have been improved for better field persistence[45][46]. To address these issues, modern formulations include UV protectants, sticking agents, or microencapsulation to prolong the efficacy of Bt in the field[45]. Another consideration is resistance management – since Bt sprays have been used for decades, some pest populations (e.g. diamondback moth Plutella xylostella) have evolved resistance to certain Bt strains when used intensively. Rotating Bt with other biocontrol agents or chemicals is recommended to mitigate resistance development.
Nevertheless, when applied at the right time (e.g. early instar larvae) and under favorable conditions, Bt biopesticides can match or even outperform chemical insecticides[4]. They have been credited with saving certain crops from pests without the environmental toll. For example, Bt sprays are used extensively in crucifer vegetable production against leaf-eating caterpillars, reducing the need for organophosphate and pyrethroid sprays. In forestry, aerial spraying of Btk has been used to combat invasive moth outbreaks (like gypsy moth) while minimizing harm to other wildlife, a clear environmental benefit. In mosquito abatement, Bti has largely replaced chemical larvicides in many wetland areas, thus protecting aquatic ecosystems while controlling disease vectors. In summary, Bacillus thuringiensis as a microbial insecticide has proven to be a powerful tool for sustainable agriculture. Its use aligns with IPM principles: it is compatible with natural enemies, leaves no toxic residues, and can be used in rotation or combination with other methods. The widespread adoption of Bt biopesticides in both conventional and organic systems attests to its success. Going forward, improvements in formulation technology and discovery of new strains promise to further enhance the reliability of Bt products[45][46].at embracing such biocontrol agents is an investment in the future – as regulatory pressures and resistance issues limit fungicides, having a reliable bio-tool like Ampelomyces is invaluable for sustainable agriculture.
Genetically Modified Bt Crops#
Perhaps the most transformative application of B. thuringiensis in agriculture has been the development of Bt crops – plants genetically engineered to express Bt toxin genes. Bt crops bring the insecticidal power of Bt into the plant itself, providing season-long protection against target pests without the need for external spray applications. This technology was a breakthrough in the mid-1990s and has since been widely adopted in many parts of the world.
The first Bt crop to be commercialized was Bt potato (NewLeaf™), approved in 1995 in the United States to control Colorado potato beetle. However, the Bt potato was relatively short-lived on the market. The major impact came with Bt cotton and Bt maize (corn), both introduced in 1996. Bt cotton was engineered to control lepidopteran bollworms (such as the pink bollworm and cotton bollworm), which are devastating pests in cotton production. Bt maize was initially designed to control the European corn borer and related stem-boring caterpillars; later versions also targeted corn rootworms (beetles) by using different cry genes. Since their introduction, Bt cotton and Bt maize have been adopted on tens of millions of hectares globally, making them among the most rapidly adopted crop technologies in modern agriculture[47][48].
Global adoption: Over the past two decades, Bt crop adoption has grown dramatically (Table 2). As of 2019, genetically modified (GM) maize expressing Bt toxin was grown on about 60.9 million hectares worldwide, representing roughly 32% of global maize area[49]. Likewise, Bt cotton covered approximately 25.7 million hectares (about 14% of global cotton area in 2019)[49]. Major countries growing Bt maize include the United States, Brazil, Argentina, South Africa, Canada, and the Philippines, among others. Major Bt cotton producers include India, China, the United States, Pakistan, Brazil, and Australia. Notably, India’s cotton farming underwent a revolution after Bt cotton was introduced in 2002 – by 2017, around 95% of India’s cotton area (over 11 million hectares) was planted to Bt cotton hybrids[50]. China also rapidly adopted Bt cotton in the late 1990s, virtually eliminating severe bollworm outbreaks that once plagued its cotton fields. In the United States, over 80% of cotton and ~80% of maize acreage is now planted to varieties with Bt traits (often combined with herbicide-tolerance traits). Bt soybean (with Cry1Ac to control caterpillars) has been grown in South America (Brazil, Argentina) since 2013, and Bt brinjal (eggplant) was commercialized in Bangladesh in 2013—the first Bt vegetable crop, targeting the fruit-and-shoot borer. Bt technology has even been applied to other crops like Bt rice (approved in China in 2020 for pest-resistant rice, though not yet widely grown) and Bt cowpea (approved in Nigeria in 2019 to control pod-borer in cowpeas). These developments illustrate the expanding scope of Bt crops beyond the initial cotton and maize.
Table 2. Global Adoption and Effectiveness of Key Bt Crops (2000s–2020s)
| Bt Crop (Target pests) | First Release | Major Adopting Regions | Adoption Scale (c. 2020) | Documented Benefits |
| Bt Maize (corn)<br/>Targets: Stem borers (Ostrinia, Sesamia), earworms, rootworms (in stacked varieties). | 1996 (USA)[47]; shortly after in Canada, Argentina, S. Africa. | Americas (USA, Brazil, Argentina, Canada), Asia (China, Philippines), Africa (S. Africa, Nigeria in trials). | ~60 million ha globally (2019)[49] (~32% of world maize area). In US/Brazil, >80% of maize contains Bt trait (often stacked). | ~+5–10% yield increase in regions with high pest pressure; reduced insecticide sprays (e.g. less need for corn borer spraying)[51]. Regional pest suppression observed (e.g. area-wide reduction of European corn borer)[52]. Improved grain quality (less damage and mycotoxin). |
| Bt Cotton<br/>Targets: Bollworms (Helicoverpa/Heliothis, Pectinophora), budworms and armyworms. | 1996 (USA); 1997 (China); 2002 (India)[53]. | Asia (India, China, Pakistan), Americas (USA, Brazil, Argentina), Africa (South Africa, Sudan, Malawi), Australia. | ~25 million ha globally (2019)[49] (~14% of world cotton area). Near-saturation adoption in India (~95%)[50] and China (~90%+); high adoption in USA (>80%). | Significant yield gains (up to +20% in smallholder systems) due to less boll damage[51]. Major reductions in chemical insecticide use (50% or more fewer sprays for bollworms)[54], translating to cost savings and health benefits for farmers. Enhanced beneficial insect activity and easier IPM of secondary pests in some cases[39][40]. |
| Bt Eggplant (Brinjal)<br/>Target: Eggplant fruit and shoot borer (Leucinodes orbonalis). | 2013 (Bangladesh)[55]; 2021 (Philippines). | South Asia (Bangladesh), Southeast Asia (Philippines); trials in India (not yet approved). | ~1,500–2,000 ha in Bangladesh by smallholders (as of late 2010s), growing yearly[56]. Still limited but expanding adoption. | Nearly eliminates pest damage (up to 95% reduction in fruit infestation reported). Up to 80% fewer insecticide applications on Bt brinjal fields[54], improving farm safety and profitability. Yields increased ~30–50% for Bt adopters vs. non-Bt[57]. |
Sources: ISAAA reports; Turnbull et al. (2021)[49]; Klümper & Qaim (2014) meta-analysis[51]; Bangladesh Bt brinjal study[54][57]; Tabashnik (2023); etc.
The effectiveness of Bt crops has been well documented. By having the plant produce the Bt toxin in its tissues, protection is continuous and not weather-dependent. This has led to significant decreases in crop losses due to target pests. For example, Bt cotton effectively controls the cotton bollworm complex, which in turn has raised cotton yields in countries like India and China by ~10–20% on average (and much more in seasons of heavy pest pressure)[51]. Bt maize has virtually eliminated major damage from European corn borer in North America – so much so that even non-Bt maize fields benefited from regional pest suppression, as the pest population fell due to widespread planting of Bt maize (a phenomenon known as the “halo effect” or area-wide suppression)[52]. Perhaps most impressively from an environmental standpoint, the adoption of Bt crops has sharply reduced the need for chemical insecticide sprays on those crops. A global meta-analysis of 147 studies found that, on average, farmers planting insect-resistant GM crops (mostly Bt cotton and maize) used 37% less chemical insecticide (by volume) compared to conventional crops, while achieving higher yields[51][58]. In countries like China, Bt cotton adoption led to dramatic drops in spraying for bollworms, which also resulted in increased populations of beneficial insects (like ladybird beetles and lacewings that help control other pests) in cotton fields[39][40]. In Bangladesh, Bt brinjal farmers were able to cut insecticide sprays from 25-30 times per season to 2-3 times, virtually eliminating exposure to toxic pesticides for themselves and consumers[54].
While Bt crops are a triumph for pest management, they are not a panacea and must be managed properly. In particular, the next sections will discuss insect resistance and other challenges that have emerged. However, from a sustainable agriculture perspective, Bt crops have delivered clear benefits: higher effective yields per hectare (thus helping food production without expanding land area), reduced chemical pesticide load in the environment, and compatibility with conservation of non-target biodiversity[39][40]. These outcomes align strongly with the goals of sustainable agriculture, making Bt crops a prominent success story in agricultural biotechnology.
Ecological and Environmental Safety Profile of Bt#
A critical aspect of Bacillus thuringiensis’s reputation as a “green” solution is its ecological safety. Unlike broad-spectrum chemical insecticides that can harm a wide range of organisms, Bt’s toxicity is highly specific to certain insects. Non-target organisms – including beneficial insects, wildlife, and humans – are largely unaffected by Bt, due to the specificity of the mode of action described earlier. Over decades of use, a substantial body of research has evaluated the environmental impacts of both Bt sprays and Bt crops. The consensus from these studies is that Bt is remarkably safe when used as intended, though with some caveats and the need for ongoing monitoring[39][40].
Specificity to Non-Targets#
Bt’s specificity stems from the requirement that the target organism have the right gut conditions (alkaline pH, proteases) and receptor molecules for the toxin. Most non-target animals, such as mammals, birds, fish, and even most beneficial insects, do not have these and thus are not affected. For instance, humans and other vertebrates have acidic stomachs and no Bt toxin receptors, so any Bt protein ingested is rapidly degraded and rendered harmless[40]. This is why Bt has an excellent safety record for human health: Bt formulations have been used on organic produce that is directly consumed, and regulatory assessments have consistently found no toxicity or allergenicity concerns with Bt proteins in GM crops (Cry proteins are digestible and do not resemble known allergens). Indeed, Bt is so safe to humans that some Bt products are registered for household use against mosquito larvae in ponds, etc., with no need for protective equipment beyond basic handling hygiene.
Beneficial insects such as pollinators (honeybees, bumblebees), predators (ladybugs, lacewings), and parasitoids are generally unaffected by Bt at field application rates. Numerous studies and reviews have concluded that Bt crops have no adverse effects on the abundance of non-target arthropods in crop fields, especially when compared to the insecticide-sprayed alternatives[39][40]. For example, a 2020 meta-analysis focusing on Bt crop impacts on soil invertebrates found no significant differences in soil fauna populations between Bt and non-Bt fields, reinforcing that Bt traits are not harming earthworms, springtails, nematodes, or other soil organisms[39][40]. Similarly, studies on Bt crops have shown no harm to pollinators like bees; Bt proteins (Cry1, Cry2, etc.) either are not present in nectar/pollen at significant levels or do not bind to bee gut receptors.
However, Bt is not entirely without any non-target interactions. A notable early controversy arose in 1999 when a laboratory study indicated that pollen from Bt corn (expressing Cry1Ab) could harm monarch butterfly caterpillars if the pollen were dusted on milkweed leaves (the monarch’s host plant). This study led to public concern, but subsequent extensive field research found that the risk to monarchs in the field is exceedingly low – Bt pollen deposition on milkweed in cornfields is usually below harmful levels and monarch larvae exposure is limited. The monarch case is often cited to demonstrate that while Bt is generally safe, each case should be examined: in this instance, regulators and scientists conducted the necessary research and confirmed that Bt corn posed minimal threat to monarch butterfly populations (especially relative to other factors or the alternative scenario of insecticide use)[59][60].
Another area of environmental fate is the persistence of Bt toxins in soil and water. Bt spores are natural soil inhabitants (Bt is found globally in soils even where not applied), and Cry proteins released into the environment can bind to soil particles. Studies show that Bt toxins in plant residues can persist for some weeks or months in soil, but they are eventually degraded by microbes and environmental factors[61]. Importantly, no accumulation to harmful levels has been observed under normal field conditions[62][60]. The Cry proteins bind to clays and humic substances, which reduces their bioavailability. Additionally, multiple studies (including the meta-analysis above) indicate that these residues do not negatively impact soil ecosystem services or nutrient cycling[39][40].
Aquatic environments have also been investigated, particularly for Bt israelensis used in mosquito control (which is directly applied to water) and for potential runoff of Bt proteins from Bt crop fields. Bt israelensis products have been used for decades in larvicide programs with little to no effect on non-target aquatic insects, aside from the intended mosquito and blackfly larvae[60][63]. Some sensitive aquatic invertebrates (like certain midges or micro-crustaceans) could be indirectly affected if their food web is altered (for instance, if mosquito larvae they prey on are eliminated, predators might need to adjust)[64][65]. But these are second-order ecological effects rather than direct toxicity, and overall, Bti is considered an environmentally safe mosquito control agent. For Bt crops, experiments where high levels of Bt corn byproducts were added to streams showed negligible direct effects on aquatic insects[66][67]. In real-world scenarios, any Bt protein in runoff is highly diluted.
Ecosystem and Food-Web Considerations#
While direct toxicity of Bt to non-targets is minimal, scientists have considered broader ecosystem interactions. For example, if Bt controls a pest very effectively, the predators and parasitoids that normally feed on that pest might have less food and could decline in abundance. This is an indirect effect – generally a positive one if the pest is the target (as crop damage is reduced), but it could impact food webs. In practice, reductions in broad-spectrum insecticide use associated with Bt crops often lead to increases in natural enemy populations, which likely compensates for any reductions due to fewer pests[68][40]. In Bt cotton fields in China, for instance, generalist predators thrived because farmers sprayed much less insecticide for bollworms, resulting in better biological control of other pests like aphids by those predators (a side benefit of Bt)[39]. Thus, Bt crops can indirectly enhance biological control of secondary pests by preserving natural enemies that would have been harmed by insecticides.
Nonetheless, researchers have noted that Bt is not entirely benign in all contexts. A comprehensive review by Belousova et al. (2021) highlighted that Bt strains introduced to an ecosystem (such as through repeated spraying) could potentially interact with native microbes and even exchange genes, given Bt’s close relation to B. cereus. The authors pointed out rare cases where B. thuringiensis can carry virulence factors (like those in B. cereus) that might affect non-insect hosts[69][70]. These are mostly hypothetical or laboratory-demonstrated possibilities; in practice, no outbreaks of disease in wildlife or humans have been attributed to Bt use. The review also emphasized examining multi-trophic effects – for example, if Bt kills caterpillars, insectivorous birds might have less prey, though this is usually minor compared to the environmental benefits of reduced chemical use[71][72]. Overall, the recommendation is that Bt-based pest control should continue to be evaluated in a holistic ecosystem context, but thus far it has proven to be one of the most environmentally selective pest control methods available.
Human and Animal Safety#
From a human health perspective, Bacillus thuringiensis has an exemplary safety record. Bt sprays have been used on commercial organic produce right up to harvest. The bacterial spores can be present on food, but there is no evidence of pathogenicity; our digestive conditions prevent Bt from causing any infection or illness. Regulatory agencies have set tolerances exempting Bt residues on food because of its demonstrated safety. Some people may exhibit mild skin or eye irritation if directly exposed to concentrated Bt formulations (as with any particulate material), but these are rare and easily mitigated with protective clothing during application.
For Bt crops, extensive food and feed safety assessments have been done for each introduced Cry protein. These include digestibility tests (showing the proteins break down rapidly in gastric fluid), heat stability tests, and checking for any similarity to known allergens or toxins. Cry1Ac, Cry2Ab, Cry3Bb, Vip3A, etc., all have been approved as safe in numerous countries. Livestock feed studies (e.g., feeding Bt corn or Bt cottonseed to animals) have shown no difference in animal health or product composition (milk, meat, eggs) compared to non-Bt feed. Furthermore, after over 20 years of human consumption of Bt maize and Bt cotton oil (and animals consuming Bt grain and forage), there have been no verifiable reports of adverse health effects[73]. This track record has helped build a scientific consensus that Bt crops are as safe as their conventional counterparts.
One area that initially caused concern was whether the Bt toxin gene or protein could affect gut microflora or transfer to microbes (horizontal gene transfer). Studies have not found any functional transfer of cry genes from transgenic plants to gut microbes in animals or humans. The proteins themselves, if ingested, are destroyed by digestion and thus do not persist in the gastrointestinal tract.
In conclusion, Bacillus thuringiensis’s environmental and safety profile is a major reason it is heralded as a green solution. Bt targets pests with precision. By doing so, it reduces collateral damage to ecosystems compared to chemical pesticides, which often kill indiscriminately. The use of Bt, especially in Bt crops, has been correlated with higher on-farm biodiversity (insects, birds) relative to sprayed fields, and meta-analyses reinforce that Bt causes no significant harm to non-target species at the community level[39][40]. Continued stewardship – such as monitoring resistance (discussed next) and ensuring balanced IPM approaches – will help maintain these benefits long-term.
Emerging Challenges: Insect Resistance and Regulatory Concerns#
Insect Resistance to Bt Toxins#
The evolutionary Achilles’ heel of any pest control strategy is the potential for the target pests to develop resistance. Just as insects have evolved resistance to many chemical insecticides, they can also evolve resistance to Bt toxins if exposed over many generations. In fact, the very specificity and potency of Bt can create strong selective pressure on pest populations in Bt crop settings. Since the late 1990s, resistance management has been a central component of Bt crop deployment, and it remains a critical challenge moving forward[74][75].
Documented cases of resistance: In the initial years of Bt crop use, field failures due to resistance were rare. High-dose strategies and refuge requirements (planting a fraction of acreage with non-Bt plants to harbor susceptible insects) were effective at delaying resistance. However, with over 25 years of Bt crop cultivation, some pest populations have evolved significant resistance. A 2023 global review by Tabashnik et al. found that by 2020, practical resistance (field-evolved resistance that reduces Bt crop efficacy) had been documented in populations of 11 pest species across 7 countries, involving 9 different Bt toxins used in crops[76]. This was a jump from just a few cases in the early 2000s. Notable examples include:
- Pink bollworm (Pectinophora gossypiella) – developed resistance to Cry1Ac in Bt cotton in India by the late 2000s, leading to control failures. (In the U.S., resistance was largely averted by mandating refuges and later by switching to two-toxin cotton, after which pink bollworm was actually eradicated regionally.)
- Cotton bollworm (Helicoverpa zea, also called corn earworm) – in parts of the southern United States, this pest evolved resistance to Cry1 and Cry2 toxins in Bt cotton by around 2014, resulting in increased damage; it is now managed with a third toxin (Vip3A) in newer varieties[77].
- Fall armyworm (Spodoptera frugiperda) – in Brazil and some African countries, resistance to Cry1F maize was observed within 10 years of deployment, reducing its effectiveness.
- Western corn rootworm (Diabrotica virgifera virgifera) – notable as a beetle rather than a caterpillar, it evolved resistance to Cry3Bb1 and related rootworm-targeting Bt proteins in parts of the U.S. Corn Belt, due to continuous corn planting and insufficient rotation/refuge.
- Diamondback moth (Plutella xylostella) – a pest of crucifer vegetables, which developed resistance to Bt sprays in some regions as early as the 1990s, and is a cautionary tale for overuse of a single Bt strain in foliar applications.
Field resistance generally manifests as higher survival of larvae on Bt plants, leading to unexpected damage. When such cases emerge, the typical response is to implement mitigation: for instance, switching to a different Bt toxin (if available), increasing refuge size, or using integrated measures (supplemental insecticides or biological controls) for a time. The good news is that many pests remain susceptible. The same 2023 review noted that for 16 major pest species in 10 countries, there was no significant decrease in susceptibility to Bt crops even after up to 20 years of exposure[78]. This demonstrates that resistance is not inevitable if managed well.
Mechanisms of resistance: Insects can resist Bt toxins through several mechanisms: changes in gut binding sites (receptor mutations or downregulation are common – if the toxin can’t bind, it can’t form pores[75]), increased immune response or midgut regeneration, behavioral avoidance (e.g. feeding less on Bt tissues), or metabolic processes that degrade the toxin. The most common are mutations in genes encoding the receptor proteins (such as cadherin or aminopeptidases) that Cry toxins bind to. These have been found in pink bollworm and others. Some insects have also shown changes in gut proteases that result in incomplete activation or faster degradation of the toxin. Interestingly, many field-evolved resistances are recessive at the genetic level (meaning an insect needs two copies of the resistance allele to survive on Bt). This fact was anticipated and underpins the refuge strategy: by ensuring that susceptible insects (with two susceptible alleles) are common, most resistant offspring (from a mating of resistant and susceptible) will be heterozygous and still get killed by the high toxin dose in Bt plants[75].
Resistance management: To prolong the efficacy of Bt, proactive insect resistance management (IRM) strategies are essential. Key approaches include:
- High Dose / Refuge strategy: Bt crops are engineered to express a high level of toxin intended to kill >99.9% of heterozygous resistant larvae, so that only truly resistant homozygotes survive (which are extremely rare if resistance alleles are rare). Non-Bt refuges (a certain percentage of crop area planted to non-Bt varieties) produce abundant susceptible insects that mate with any rare resistant survivors, making it unlikely for two resistant alleles to pair up[75]. This strategy was mandatory in the U.S. and has been credited with delaying resistance in pests like European corn borer and tobacco budworm.
- Pyramiding toxins: Next-generation Bt crops often contain two or more different Bt toxin genes targeting the same pest. If the toxins have different modes of binding, an insect resistant to one will likely be killed by the other (“redundant killing”). Examples: Bollgard II cotton (Cry1Ac + Cry2Ab), VT Double/Triple Pro corn (Cry1A + Cry2Ab for caterpillars; plus Cry3Bb for rootworms in triple). Pyramids can dramatically improve durability, especially if the chance of one insect having resistance to both toxins is extremely low. Field data support that pests took longer to adapt to two-toxin crops than single-toxin ones[79][74]. However, if cross-resistance occurs or one toxin is much weaker, the pyramid may function like a single toxin. Newer pyramids even include Vip3A (with Cry1 and Cry2) for lepidopterans and a combination of Cry3, Cry34/35, and now RNAi for rootworms in maize.
- Crop/trait rotation: Rotating different Bt toxins in time (one season use Bt toxin A, next season toxin B) could delay resistance by not giving continuous selection to one toxin. In practice, this is harder to implement because seed choices are driven by market and companies often stack all traits together. However, crop rotation (e.g. corn to soybean) inherently gives a break for pests like rootworm that only feed on corn.
- Monitoring and IPM: Regular monitoring of pest populations for early signs of resistance (using field survival assays or diagnostic doses in lab) allows for responsive management. If resistance is detected early, authorities can recommend integrated pest management steps – such as using pheromone traps, biological pesticides (like spraying Bt or virus sprays to kill survivors), or even limited use of conventional insecticides in hotspots – to knock down resistant populations. Integrating natural enemies can also help, as some resistant insects may have fitness costs that predators or parasitoids exploit.
- Next-gen technologies: Researchers are exploring novel methods like gene editing of pest populations, or gene drives, and RNA interference (RNAi) where plants produce RNA that silences critical pest genes. Notably, the latest product for corn rootworm control combines Bt and RNAi (a trait that produces a RNA targeting a rootworm gene): the idea is that even if rootworms resist Bt, the RNAi will kill them, and vice versa. These combined approaches are seen as future tools to manage resistance.
The evolution of resistance is an ongoing “arms race” between pest genetics and human management. Bt remains effective against most target pests, but vigilance is needed. The record so far shows that where robust IRM was practiced (e.g., in the U.S. with structured refuges), resistance took longer or hasn’t occurred for some pests[76][75]. Conversely, in regions with inadequate refuge compliance or where single-toxin products were used extensively without alternation (e.g. parts of India, or early Bt corn in Puerto Rico for fall armyworm), resistance can appear in a matter of years. The lesson is clear: to preserve the sustainability of Bt as a resource, proactive resistance management is indispensable.
Regulatory and Societal Concerns#
The rise of Bt, especially in the form of GM crops, has not been without controversy. While the scientific community largely agrees on the safety and benefits of Bt technologies, public perception and regulatory policies have varied widely around the world[80]. Two main areas of concern are often cited: environmental and food safety (addressed in the previous section, but still a public discussion point) and socio-economic/regulatory issues including labeling, trade, and ethics of GM crops.
Regulatory landscape: The use of Bt sprays is generally well accepted and regulated similarly to other biopesticides. Because Bt is a naturally occurring organism and has a long safety record, most countries allow Bt biopesticides even in sensitive habitats and organic farming. Regulatory oversight focuses on quality control (ensuring formulations indeed contain only the intended Bt strain and do not have contaminants) and on application guidelines. There is little controversy in this arena; Bt biopesticides have been a positive story of a safe pest control agent.
In contrast, Bt crops (GMOs) face divergent regulatory treatment. The United States, Canada, Brazil, Argentina, Australia, India, China, Philippines, Bangladesh and a number of other countries have approved and widely adopted Bt crop technology. On the other hand, many European countries, as well as some in Africa and Asia, have restrictions or bans on cultivation of GM crops, including Bt crops[80]. The European Union has a rigorous regulatory process and historically a cautious stance: as of 2025, only one Bt maize (MON810) was ever grown to some extent in Europe (notably Spain), and several EU nations disallowed even that. The reasons are often less about scientific risk – the European Food Safety Authority has repeatedly found Bt crops safe – and more about public opinion, political factors, and the precautionary principle. Concerns in these regions include long-term ecosystem effects, corporate control of seeds, and a desire to maintain a GMO-free status for market reasons. In contrast, countries like the US, Brazil, and India have embraced Bt crops as tools for farmers, with India’s example showing a massive boost in cotton production after Bt adoption.
Safety controversies and public perception: A “world controversy” about Bt crops’ safety indeed occurred, not always grounded in scientific evidence[81]. Anti-GMO advocacy groups have at times claimed that Bt toxins could harm humans or animals, or that Bt genes might unpredictably affect plant metabolism. Extensive testing and 25+ years of consumption have not borne out these claims[73]. Nevertheless, trust is a key issue – in some regions farmers or consumers have been skeptical of Bt crops, viewing them as foreign or risky. Misinformation can spread, for instance the unfounded rumor that Bt corn harmed monarch butterflies persisted even after it was debunked. Regulatory agencies thus face the challenge of communicating the science effectively. The case of Bt brinjal in India is illustrative: although field trials and science panels in India found it to be safe and beneficial, a moratorium was placed in 2010 due to activist pressure and political decisions, delaying its approval (neighboring Bangladesh went ahead in 2013 and has seen success with farmers). This shows how regulatory outcomes can hinge on public acceptance as much as on scientific data.
Another regulatory concern is the biosafety of gene flow – could Bt genes escape into wild or weedy relatives and cause problems? For example, Bt corn could potentially cross with teosinte (wild corn in Mexico) or Bt cotton with wild cotton species. If such gene flow occurred, a wild plant might gain insect resistance, possibly giving it a competitive edge (a weediness concern). In practice, most crops that have wild relatives in their centers of origin are either not yet approved for Bt traits or are managed (e.g. buffer zones). For cotton, wild Gossypium are limited geographically; for corn in Mexico, studies found some presence of transgenes in landraces early on, but no evidence of spread or weediness issues. Rice could be a concern in the future, since wild rice species exist—this will be monitored as Bt rice gets deployed.
Economic and ethical considerations: Some critics argue that Bt crops, developed by large biotech companies, can make farmers dependent on purchasing seeds annually and could erode traditional seed-saving practices. There was controversy in India around the cost of Bt cotton seeds (initially higher) and allegations that it contributed to farmer debt, although studies generally show Bt cotton increased profits overall due to yield gains and cost savings on pesticides. Intellectual property issues also arise: patents on Bt genes raised debates about access, though some public-sector Bt varieties have been developed (e.g. in India and China) to address this. From a regulatory perspective, governments have to balance enabling access to beneficial technology with appropriate safeguards and benefit-sharing.
Regulatory science: Modern regulatory evaluations for Bt crops are extensive. Before approval, data must be submitted on molecular characterization of the inserted genes, environmental risk assessment (including non-target organism testing, gene flow assessment, etc.), and food/feed safety studies. For example, feeding studies with livestock, allergenicity simulations, and environmental fate of the Bt protein are all scrutinized. In countries with established frameworks (US, EU, etc.), these processes are rigorous and involve expert review. The length and cost of regulatory approval can be very high – an oft-cited figure is that bringing a GM crop to market can take a decade and tens of millions of dollars in regulatory expenses. This has meant that primarily large companies could afford to commercialize Bt crops, which is part of the socioeconomic debate.
There are also regulatory concerns about resistance management – in the US, the EPA conditions for Bt crop approval included refuge requirements and monitoring obligations. In some other countries, enforcing such compliance is challenging (e.g. refuge planting was not well-enforced in India, contributing to pink bollworm resistance). Regulators today are considering how to update these requirements (for instance, requiring integrated seed mixes for refuges in corn, or mandating pyramided toxins only).
Finally, a positive regulatory story is that Bacillus thuringiensis has helped reduce the environmental load of more hazardous pesticides. A study on GM crop impacts from 1996–2018 estimated that insect-resistant crops reduced global insecticide active ingredient use by millions of kilograms and significantly reduced the environmental impact of pest control on those hectares[82]. This aligns with international goals to reduce chemical pesticide use for sustainability.
In summary, the challenges on the regulatory and public front are not about the efficacy of Bt – which is well proven – but about acceptance and governance. Continued transparent research, monitoring, and communication are vital. The concept of sustainable agriculture includes not only environmental and economic factors but also social ones. Thus, addressing the valid concerns of stakeholders, ensuring smallholder farmers can benefit from Bt (as seen in countries like Bangladesh with Bt eggplant), and tailoring regulations to local contexts are all important for the future of Bt technology. As the world faces increasing food security pressures and a need for eco-friendly farming, building trust in technologies like Bt is crucial. Over time, as evidence accumulates and new generations of consumers and farmers emerge, Bt may gain broader acceptance, much as past innovations eventually did.
Innovations and Future Prospects in Bt Biotechnology#
Even as Bacillus thuringiensis is firmly established in today’s agricultural practices, research and innovation continue to expand its potential. Several avenues of advancement can be identified: discovery of new toxins and enhancement of existing ones, novel formulation and delivery methods, integration with other biotechnologies (like gene editing or RNAi), and diversification of Bt’s applications beyond insect control (such as disease control or plant growth promotion). Here we discuss some of the key future prospects for Bt as a sustainable agriculture ally.
New Toxins and Engineering for Broader Pest Control#
One area of active research is the search for new Bt toxins that can target pests which current Cry/Vip proteins do not effectively control. For instance, hemipteran pests (like aphids, whiteflies) and certain nematodes cause major crop damage but are generally not susceptible to known Bt toxins. Scientists are mining the vast genetic diversity of B. thuringiensis and related Bacillus species for novel toxins. Advances in genomic sequencing have led to identification of many new cry and vip genes in Bt strains from various environments. Some have unique specificities – for example, recently characterized toxins in Cry7, Cry8, Cry51 etc. with activity on pests like plant bugs or locusts. While none of these are commercial yet, in the future we might see Bt genes conferring resistance to sap-sucking pests, which would be revolutionary for crops like citrus, grapes, or soybeans where such pests are now controlled only by chemical sprays.
In parallel, protein engineering is being applied to Bt toxins. Researchers have experimented with domain swapping or site-directed mutagenesis of Cry proteins to alter their properties – such as widening their host range or overcoming insect resistance. For example, modifying a toxin’s binding domain to bind a different receptor could retarget it to a new insect order. Another strategy is to create chimeric toxins that combine the lethal traits of two toxins. Although this is complex, it has yielded some success in the lab, and one day engineered “smart toxins” could be deployed if they prove safe and effective.
It’s worth noting that Cry1Ac and Cry2Ab – mainstay toxins in many Bt crops – are decades old in terms of discovery. New variants, like Cry1F, Cry1Ae, Cry1Ie, have supplemented the arsenal, and Vip3A (not a Cry but often classed among Bt toxins) has become a crucial component in cotton and corn due to its different mode of action which can kill certain Cry-resistant insects[83]. Going forward, stacking multiple Bt genes will remain a primary strategy to ensure robust pest control and delay resistance. Already, the trend is towards three-gene or four-gene stacks in a single plant (e.g., SmartStax corn has six insecticidal genes: multiple Cry’s for above-ground pests plus Cry34/35 for rootworm and now an RNAi for rootworm too). This stacking could extend to incorporate non-Bt insecticidal proteins as well, such as plant-derived toxins or other bacterial toxins, creating a broader spectrum bio-control crop.
Improved Formulations and Delivery#
For Bt biopesticide products, formulation technology is an ongoing area of innovation. Researchers are developing formulations that improve shelf-life, field persistence, and efficacy of Bt. Examples include microencapsulation (encasing Bt spores/crystals in a protective polymer that releases them slowly and shields from UV), wettable granules for better soil application, and oil-based formulations for better adherence to leaves. Encapsulation can also allow combination of Bt with other agents (like feeding attractants or other biopesticides) to increase effectiveness. Some studies are exploring nano-formulations, where Bt toxins are delivered via nanoparticles, potentially enhancing stability or allowing targeted delivery on a microscopic scale.
Another frontier is transformation of other organisms with Bt genes to serve as delivery vehicles. For instance, engineering endophytic fungi or bacteria that live within plants to produce Bt toxins could give plants inherent protection without being a GMO plant themselves. There have been experiments with Pseudomonas fluorescens bacteria engineered to carry cry genes, which can colonize plant roots and provide insect resistance[84]. While not yet commercial, this approach might be useful especially in regions averse to GM crops but open to biocontrol microbes.
Sprayable Bt toxin proteins (Bioinsecticides): One innovation already on the market is the production of purified or recombinant Bt toxins (rather than whole bacteria) for spray use. This can allow delivering high concentrations of specific Cry proteins, possibly including those not easily produced in large quantities by Bt fermentation. Recombinant production (e.g., in fermenters using other bacteria or in plant cell culture) might yield more cost-effective or tailored Bt products, and since it’s just protein, regulatory hurdles might differ (though cost is a factor).
Integration with Other Biotechnologies#
Bt technology is increasingly being combined with other advanced approaches to pest management:
- RNA interference (RNAi): As mentioned, RNAi traits are being stacked with Bt to attack pests through a second, unrelated mechanism (gene silencing). This is seen as a powerful one-two punch, particularly for pests like corn rootworm where RNAi can silence essential genes and Bt covers the rest[85]. In foliar pests, future Bt plants might come with both Bt toxins and RNAi targeting, say, a detoxification gene of the pest, making the pest hyper-susceptible to the toxin.
- Gene drives and Sterile Insect Techniques: In principle, gene drive systems (not yet deployed in agriculture) could spread susceptibility alleles or otherwise make pest populations more controllable. While not directly involving Bt, a wild idea is a gene drive that makes pests re-sensitive to Bt toxins if they had developed resistance. These are speculative but being discussed as possible complements to Bt.
- Genome editing in crops: CRISPR-based genome editing is enabling new forms of pest-resistant crops that are not transgenic (no foreign gene) but edited for traits like producing insect-deterring compounds. While different from Bt, they could be used together: e.g., an edited plant that produces a defense metabolite plus a Bt toxin gene for dual protection. Moreover, CRISPR might be used to create plants that express Bt genes in more targeted ways (like only in certain tissues or at certain times) to reduce selection pressure – although current transgenics already use tissue-specific promoters to some extent (e.g., root-specific expression for rootworm).
- Synthetic biology: There is exploration of using synthetic biology to modify Bacillus thuringiensis strains themselves. For instance, one could engineer a single Bt strain with an expanded toxin portfolio (covering multiple pest orders) to use as a super-bioinsecticide. Or engineering Bt to be a more robust plant colonizer (combining traits of Bt with plant growth promoting bacteria) so that when applied, it not only kills pests but also establishes a beneficial presence on the plant.
Beyond Insect Control: Bt as a “Green Microbe” in Other Roles#
In line with the concept of Bt as a green microbe, researchers have uncovered roles for Bacillus thuringiensis beyond entomopathogenic activity. Some Bt strains have shown antimicrobial activity against plant pathogens, producing compounds like zwittermicin (an antibiotic) that can suppress certain crop diseases. This raises the prospect of Bt-based treatments for fungal or bacterial plant diseases, either through foliar sprays or soil inoculants[86]. Bt is a close relative of Bacillus cereus and Bacillus subtilis, some of which are known biocontrol agents against diseases; thus, it’s not surprising that Bt too has such potential if the right strain or metabolite is identified.
Another exciting area is plant growth promotion. Several studies have found that Bt is capable of colonizing the rhizosphere (root zone) or even endophytic niches in plants and can act as a plant growth-promoting rhizobacterium (PGPR)[87][88]. For example, certain Bt strains can solubilize insoluble phosphates in soil, making phosphorus more available to plants, similar to how some soil bacteria promote fertility[89]. Others produce siderophores, molecules that chelate iron and help plants acquire this nutrient[90]. Experiments co-inoculating Bt with legume symbionts have shown increased nodulation and plant biomass in soybean and peas[91][92]. One particular Bt strain (Bt NEB17) produces a compound called Thuricin 17, a bacteriocin that at low concentrations can stimulate plant root growth and stress tolerance[93]. This is being investigated as a bio-stimulant to enhance crop vigor and yield (e.g., under drought conditions, Thuricin 17-treated plants showed improved performance). The idea that Bt could double as both an insect defense and a growth-promoter is quite appealing for sustainable agriculture, potentially reducing the need for chemical fertilizers and pesticides simultaneously.
Bt has also found some industrial and environmental applications. For instance, Bt can produce polyhydroxyalkanoates (bioplastics precursors) under certain conditions[94]. Some strains have been studied for bioremediation, such as breaking down or immobilizing heavy metals in contaminated soils[94]. There is even research into using Bt or its spores in novel ways like nanoparticle synthesis for agricultural or medical use[95]. Moreover, the Parasporin proteins from Bt, which are not insecticidal, have shown selective toxicity against cancer cells, hinting at possible biomedical applications of Bt-derived proteins[86].
Prospects and Outlook#
Looking ahead, Bacillus thuringiensis is likely to remain at the forefront of biological pest management, continually evolving with the help of science. In the next decade or two, we might see:
- Next-gen Bt crops that incorporate multiple modes of action (stacked traits including Bt toxins, RNAi, and perhaps novel traits) to counter pest resistance and target a broader pest complex.
- Expansion of Bt crop varieties: More crops engineered with Bt genes (subject to regulatory approval and public acceptance). For example, Bt eggplant’s success could pave the way for other vegetables, Bt cowpea in Africa is a milestone for subsistence crops, and Bt rice could benefit rice-growing regions if deployed cautiously to avoid resistance.
- Refined deployment strategies: For instance, use of spatially precise application of Bt (like soil inoculation at planting with Bt for root pests, drones applying Bt sprays only when pest thresholds are met, etc.) to maximize effect and minimize selection pressure.
- Microbiome approaches: Harnessing the plant microbiome so that endophytes or symbionts carry insecticidal traits. There’s research on symbiotic bacteria in insects (like Wolbachia or gut flora) that could be engineered to deliver toxins specifically inside pests – a kind of Trojan horse approach.
- Integration into climate-resilient farming: As climate change alters pest patterns, Bt’s role might become even more vital. Warmer climates might expand ranges of certain pests; Bt crops and biopesticides can be pivotal in responding without resorting to heavier chemical use. Additionally, because Bt sprays and traits help reduce crop losses, they indirectly contribute to resource-use efficiency (more yield per input) and thus could help climate adaptation by bolstering food security.
However, these prospects will only be fully realized if the challenges discussed are addressed: resistance management must keep pace, and public/regulatory acceptance must be earned through transparency and demonstrated benefits. There is encouraging movement in some of these areas. For example, newer regulatory frameworks (in countries like Nigeria, Bangladesh) are enabling small-scale farmers to access Bt crop technology in locally important crops, which helps directly show farmers the benefits. Education on Bt safety and its environmental advantages (like the meta-analyses showing no harm to soil or non-targets)[39] will hopefully alleviate public concerns over time.
In conclusion, Bacillus thuringiensis stands as a paradigm of a “green” microbial technology that has successfully transitioned from nature to widespread agricultural use. Its continued evolution through scientific innovation promises to keep it at the heart of sustainable pest management for the foreseeable future. By learning from past experiences and embracing new knowledge, we can harness Bt in even more effective and responsible ways – protecting crops, reducing chemical inputs, and supporting the goal of a more sustainable and secure agriculture.
Summary Table: Spotlight Study#
| Category | Details |
| Lead Researchers | Lauren Huth; Gavin J. Ash; Alexander Idnurm; Levente Kiss; Niloofar Vaghefi – a team of plant pathology and mycology researchers. |
| Affiliations | Centre for Crop Health, University of Southern Queensland, Australia; School of BioSciences, University of Melbourne, Australia (among others). |
| Research Focus | Sequencing and analyzing the genome of Ampelomyces quisqualis to understand its genetic make-up, evolutionary origin, and the genomic basis of its mycoparasitic lifestyle. |
| Key Breakthroughs | – Produced the first high-quality genome assembly of A. quisqualis (≈40.38 Mb, 24 scaffolds, 96% complete)[5]. – Discovered a “bipartite” genome structure: gene-rich, GC-normal regions interspersed with large AT-rich, repeat-laden regions. Such a two-speed genome architecture is a hallmark of many plant pathogenic fungi. – Found genomic evidence suggesting Ampelomyces evolved from plant-pathogenic ancestors (shared genome features with pathogens). – Clarified that Ampelomyces consists of multiple phylogenetic lineages (at least 9 MOTUs); the sequenced strain belongs to the same clade as the commercial AQ10 strain, whereas another sequenced strain (“HMLAC 05119”) belongs to a different clade. |
| Collaborative Efforts | Involved collaboration between universities in Australia and experts in fungal genomics and pathology. The team combined expertise in biocontrol (Ash, Kiss) and molecular genetics (Idnurm, Vaghefi) to interpret the data, and referenced work by international colleagues (e.g., in Hungary, Japan) for comparative analysis. |
| Published Work | Published in Genome Biology and Evolution (Oxford University Press) in August 2021. Full reference: Huth et al., 2021, Genome Biol. Evol. 13(8): evab182. This is a peer-reviewed, high-impact journal focusing on genomic and evolutionary biology. |
| Perspective | This study provides a new perspective on Ampelomyces: rather than a simple biocontrol agent, it is a fungus with a complex genome that hints at a pathogenic past. It bridges our understanding between plant pathogens and hyperparasites, suggesting evolutionary fluidity between these roles. For biocontrol practitioners, it legitimizes Ampelomyces as a thoroughly studied organism (genetically) and paves the way for genome-informed improvements in its use. |
| Publication Date | Online on August 3, 2021 (print issue August 2021). This makes it a very recent study (as of 2021) offering up-to-date insights. |
| Location | The Ampelomyces strain sequenced (BRIP 72107) was isolated from a powdery mildew on Cestrum (ornamental plant) in Toowoomba, Queensland, Australia. The genome work was carried out in labs at University of Southern Queensland and University of Melbourne (Australia), with the strain stored in the Queensland Plant Pathology Herbarium (BRIP). The study thus had an Australian nexus, though it incorporated comparative data from strains isolated in other countries (Europe, Asia) for the phylogenetic analysis. |
| Key Findings | – A. quisqualis genome ~40.4 Mb, with a bimodal GC content (AT-rich vs GC-rich regions). About ~45–50% of the genome is repetitive/AT-rich, suggesting active transposons and a “two-speed” genome strategy. – Genomic layout and composition are similar to certain plant pathogens (e.g., Leptosphaeria maculans), supporting an evolutionary link. – Phylogenetics: The sequenced strain and the AQ10 biocontrol strain are in the same clade (MOTU1), confirming AQ10’s taxonomic placement. Another commercial strain (“Q-fect” from Korea) and a previously sequenced strain form a separate clade (MOTU4), showing cryptic speciation within Ampelomyces. – Highlighted the need for taxonomic revision: formally, all these strains are called A. quisqualis, but genetically they may be distinct species. A comprehensive polyphasic taxonomy study was suggested. – Provided a catalog of genes: identified genes for cell wall degrading enzymes, stress response, and other factors potentially important for mycoparasitism. This offers targets for future functional studies or strain enhancement. |
Conclusion#
Over the last century, Bacillus thuringiensis has transformed from a scientific curiosity into a pillar of sustainable agriculture. As chronicled in this review, its journey – from Ishiwatari’s silkworm pathogen in 1901 to the extensive Bt crop fields of today – mirrors the growing desire for eco-friendly solutions in farming. Bt’s unique insecticidal proteins offered a novel mode of action that revolutionized pest control by providing specificity and safety unimaginable with broad-spectrum chemicals of the past. This “green microbe” has enabled farmers around the world to manage pests more judiciously: Bt biopesticide formulations give organic and conventional growers a potent tool against caterpillars, beetles, and flies, while Bt transgenic crops have delivered higher yields and reduced pesticide dependency on a global scale[51][39].
Bt’s success is underscored by its environmental profile. Studies resoundingly show that Bt, whether sprayed or expressed in plants, spares non-target organisms and allows biodiversity to flourish relative to chemical-intensive practices[39][40]. This aligns perfectly with the ethos of sustainable agriculture – producing adequate food while safeguarding the agro-ecosystem. The ecological harmony is such that Bt has even been called an “environmentalist’s insecticide,” and decades of safe use validate that label. Nonetheless, this review has also addressed the need for vigilance: nature is not static, and insect populations can adapt. The emergence of Bt-resistant pest strains in some locales is a reminder that sustainability requires active management – strategies like refuge planting, toxin rotation, and pyramiding must continue to be enforced and refined[74][75]. With coordinated resistance management, the efficacy of Bt can be extended for future generations of farmers.
Regulatory and societal acceptance of Bt, especially in GM crops, has been a mixed landscape. But there are positive signs as more data accumulate and success stories emerge (for example, the rapid adoption and benefits of Bt cotton in India, or the health and yield improvements from Bt eggplant in Bangladesh[96][97]). Science-based policies and public education will be critical to overcome unfounded fears and to ensure equitable access to Bt technology, particularly for smallholder farmers who stand to gain the most from reduced crop losses and less exposure to toxic pesticides. In that sense, Bt is not just a tool for wealthier industrial farms; it can be a boon for sustainable development and food security in developing nations.
Looking to the future, Bacillus thuringiensis is poised to remain at the cutting edge of agricultural biotechnology. Innovations are expanding its utility – from new insecticidal proteins to synergy with approaches like RNAi and microbiome engineering. At the same time, research into Bt’s non-insecticidal benefits (such as plant growth promotion and disease suppression) could broaden its role as a multi-functional bioagent in crop production[89][88]. These exciting prospects reinforce that Bt is not a static technology but a dynamic one, continually shaped by scientific discovery. As climate change and global population growth intensify the pressure on agriculture, solutions that are both productive and sustainable will be in high demand. Bt exemplifies such a solution, having already contributed to increases in yield and reductions in chemical pesticide use on millions of hectares.
In conclusion, Bacillus thuringiensis has truly earned its moniker as a “green microbe.” Its advances have shown that it is possible to reconcile high agricultural productivity with environmental stewardship. The challenges that lie ahead – managing resistance, harmonizing regulations, and keeping pace with evolving pest complexes – are significant but surmountable with informed, science-driven strategies. By learning from the past and investing in future innovations, the agricultural community can ensure that Bt’s legacy continues to grow. In doing so, we not only honor a century of progress but also reaffirm our commitment to an agriculture that is both bountiful and sustainable for the planet.
References#
Abbas, M. S. T. (2018). Genetically engineered (modified) crops (Bacillus thuringiensis crops) and the world controversy on their safety. Egyptian Journal of Biological Pest Control, 28(1), 1–12.
Belousova, M. E., Malovichko, Y. V., Shikov, A. E., Nizhnikov, A. A., & Antonets, K. S. (2021). Dissecting the environmental consequences of Bacillus thuringiensis application for natural ecosystems. Toxins, 13(5), 355[10][71].
Bravo, A., Likitvivatanavong, S., Gill, S. S., & Soberón, M. (2011). Bacillus thuringiensis: A story of a successful bioinsecticide. Insect Biochemistry and Molecular Biology, 41(7), 423–431.
Chakroun, M., Banyuls, N., Bel, Y., Escriche, B., & Ferré, J. (2016). Bacterial vegetative insecticidal proteins (Vip) from entomopathogenic bacteria. Microbiology and Molecular Biology Reviews, 80(2), 329–350.
Crickmore, N., et al. (2020). Bacillus thuringiensis toxin nomenclature database (Accessed 2020)[20].
ISAAA (2019). Global Status of Commercialized Biotech/GM Crops in 2019 – Biotech Crop Annual Updates[47][98].
James, C. (2011). Global status of commercialized biotech/GM crops: 2011 (ISAAA Brief No. 43).
Jouzani, G. S., Valijanian, E., & Sharafi, R. (2017). Bacillus thuringiensis: A successful insecticide with new environmental features and tidings. Applied Microbiology and Biotechnology, 101(7), 2691–2711.
Kumar, P., Kamle, M., Borah, R., Mahato, D. K., & Sharma, B. (2021). Bacillus thuringiensis as microbial biopesticide: uses and application for sustainable agriculture. Egyptian Journal of Biological Pest Control, 31, 95[7][15].
Klümper, W., & Qaim, M. (2014). A meta-analysis of the impacts of genetically modified crops. PLoS ONE, 9(11), e111629[51].
Lambert, B., & Peferoen, M. (1992). Insecticidal promise of Bacillus thuringiensis: Facts and mysteries about a successful biopesticide. BioScience, 42(2), 112–122.
Matyjaszczyk, E. (2018). Non-chemical plant protection products – characteristics and impact on agriculture and horticulture. Journal of Plant Diseases and Protection, 125(2), 113–119.
Palma, L., Muñoz, D., Berry, C., Murillo, J., & Caballero, P. (2014). Bacillus thuringiensis toxins: an overview of their biocidal activity. Toxins, 6(12), 3296–3325.
Shelton, A. M., Zhao, J. Z., & Roush, R. T. (2002). Economic, ecological, food safety, and social consequences of the deployment of BT transgenic plants. Annual Review of Entomology, 47, 845–881[17].
Tabashnik, B. E., Fabrick, J. A., & Carrière, Y. (2023). Global patterns of insect resistance to transgenic Bt crops: The first 25 years. Journal of Economic Entomology, 116(2), 297–309[76][75].
Tripathi, V., & et al. (2020). Genetically engineered crops for sustainably enhanced food production systems. Frontiers in Sustainable Food Systems, 4, 106[49].
Vachon, V., Laprade, R., & Schwartz, J. L. (2012). Current models of the mode of action of Bacillus thuringiensis insecticidal crystal proteins: A critical review. Journal of Invertebrate Pathology, 111(1), 1–12[31].
Yang, F., Kerns, D., Head, G. P., & Huang, F. (2021). Recognition of pest resistance to Bt maize by regulatory agencies—monitoring, regulatory actions, and lessons learned. Journal of Integrated Pest Management, 12(1), 18.
Zhang, J., & et al. (2019). Comparison of the Vip3Aa protein with Vip3Aa11 and Vip3Aa39: insecticidal activity and mode of action. Pest Management Science, 75(7), 1863–1870.