Ampelomyces quisqualis: The Mycoparasitic Fungus for Sustainable Crop Protection
Overview of the Microbe Ampelomyces quisqualis is a filamentous fungus known as a mycoparasite, meaning it parasitizes other fungi. It was first described in the...
- 1
Powdery Mildew Infection on Melon Leaves and Fungal Structures

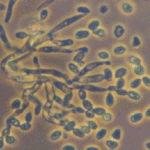
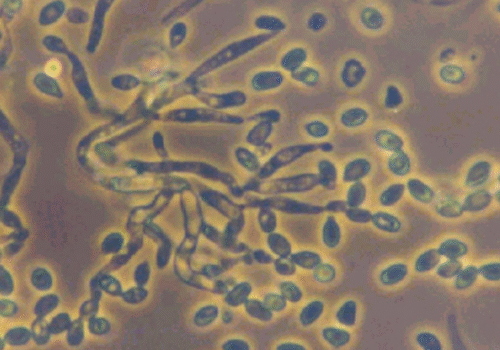
(A) Melon leaf showing white, powdery mildew patches caused by fungal growth. (B) Microscopic view of spore chains on leaf surface.
- 2
Fungal Fruiting Bodies and Spore Structures on Infected Plant Tissue
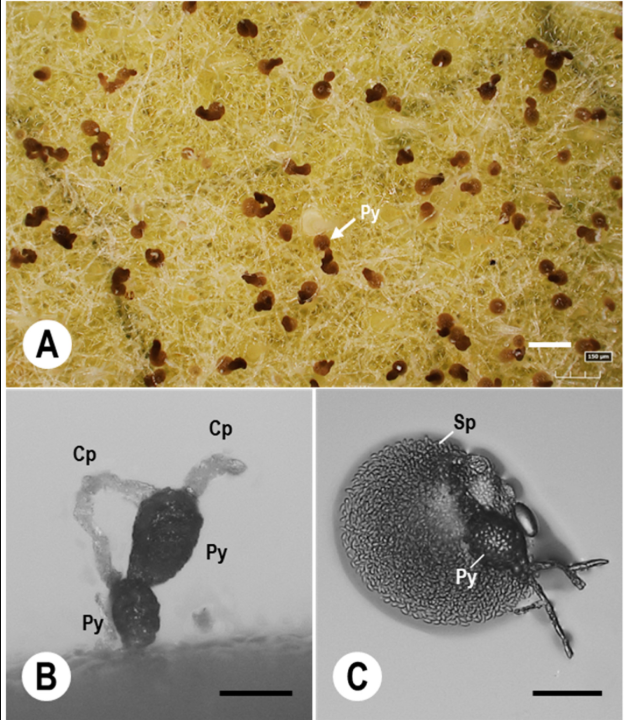
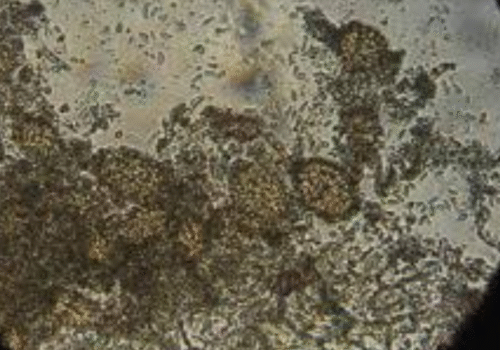
(A) Numerous dark, round fungal fruiting bodies (pycnidia) scattered across infected plant surface. (B) Close-up showing pycnidia (Py) with connecting cells (Cp). (C) Detailed view of a spore-filled pycnidium (Py) with spores (Sp) visible inside.
Overview Table of GAmpelomyces quisqualis
- Feature
Description
- Scientific Name
Ampelomyces quisqualis (Ces.) Rabenh. (1851)
- Classification
Phylum: Ascomycota; Order: Pleosporales; Family: Phaeosphaeriaceae
- Habitat
Broadly distributed as a hyperparasite on powdery mildew fungi across diverse crops and climates
- Key Functions
Mycoparasitism of Erysiphales (powdery mildew); suppression of disease spread
- Notable Abilities
Production of cell-wall–degrading enzymes; sporulation within host structures; high host specificity
- Applications
Biocontrol of powdery mildews in greenhouse and field crops; organic‐approved biofungicide formulations
- Genetic Engineering Potential
Targets: genes for chitinase and β-1,3-glucanase; tools: CRISPR/Cas9 to enhance enzyme expression
- Challenges
Variable field efficacy; humidity dependence; production scale‐up; regulatory registration
- Future Prospects
Strain improvement via synthetic biology; AI‐driven fermentation optimisation; integration in circular agrifood systems
Overview of the Microbe#
Ampelomyces quisqualis is a filamentous fungus known as a mycoparasite, meaning it parasitizes other fungi. It was first described in the mid-19th century on grapevine powdery mildew (Erysiphe necator). Historically, it was also called Cicinnobolus cesatii in older literature, but A. quisqualis is the accepted name today. Ampelomyces is widely distributed worldwide wherever powdery mildew (PM) fungi occur, and it has been found naturally infecting more than 65 species of powdery mildew fungi across at least 8 genera. This broad presence highlights its ecological role as a hyperparasite of one of the most common plant pathogen groups.
Morphologically, A. quisqualis is a pycnidial fungus – it produces flask-shaped fruiting bodies called pycnidia within the bodies of its host fungus. These pycnidia release spores (conidia) that spread to new powdery mildew colonies. Uniquely, Ampelomyces grows entirely inside its host’s fungal tissue, absorbing nutrients and eventually killing the powdery mildew. Under the microscope, its spores are hyaline (colorless), ovoid cells that form in great numbers inside the dark pycnidia embedded in the mildew’s mycelium.
In nature, A. quisqualis acts as a hyperparasite (or intracellular parasite) of powdery mildews on many plants, including important crops and ornamentals. It does not infect plants directly; instead, it targets the obligate fungal pathogens that cause powdery mildew disease on plant surfaces. This makes Ampelomyces a valuable natural ally in agricultural ecosystems, as it can suppress the growth and reproduction of powdery mildew fungi. Because chemical fungicides against powdery mildew often lead to resistance and environmental concerns, A. quisqualis has gained attention as a biocontrol agent for sustainable crop protection.
Historical note: By the early 20th century, scientists had observed Ampelomyces parasitizing powdery mildew in the field. In 1939, C.E. Yarwood reported using Ampelomyces conidia to infect and reduce mildew on plants – one of the first demonstrations of fungal biocontrol. Later, in 1977, Jarvis & Slingsby achieved greenhouse control of cucumber powdery mildew with A. quisqualis applications. These milestones set the stage for developing Ampelomyces as a practical tool in agriculture. Today, A. quisqualis is recognized as “the most studied biocontrol agent of powdery mildews” and is used in both research and commercial biopesticide products around the world.

Mycoparasitic Biocontrol of Powdery Mildews#
Powdery mildew diseases, caused by fungi in the family Erysiphaceae, affect a wide range of crops – from cereals and vegetables to grapes and ornamental plants. Ampelomyces quisqualis parasitizes a broad spectrum of these powdery mildew fungi, making it an attractive biological control agent (BCA) to combat the disease on many hosts. The biocontrol mechanism of A. quisqualis is direct and destructive: it infects the powdery mildew fungus itself, thereby protecting the plant indirectly.
Infection cycle: The process begins when Ampelomyces spores (conidia) land on a mildew-infected leaf. The conidium germinates on the plant surface and penetrates the hypha of the powdery mildew fungus. This penetration can occur through natural openings or by forming an appressorium-like structure that mechanically and enzymatically breaches the host’s cell wall. Once inside the mildew’s hypha, Ampelomyces grows internally, invading the fungal cells. It typically targets the conidiophores – the stalks of the mildew that produce its asexual spores. As Ampelomyces proliferates, it causes the mildew’s hyphae and conidiophores to atrophy and collapse. Within a few days, the powdery mildew colony’s white, fuzzy appearance diminishes as the hyperparasite destroys its host from within.
Following this internal attack, A. quisqualis undergoes sporulation: it forms new pycnidia inside the dying mildew cells. Each pycnidium produces numerous Ampelomyces conidia. When these pycnidia mature, they rupture the mildew tissue and release conidia onto the plant surface. Rain splash, wind, or simply growth of the plant can spread these conidia to other parts of the plant (or neighboring plants), where they can infect additional powdery mildew colonies. Through this repeating cycle, an initial introduction of Ampelomyces can multiply and continue seeking out new powdery mildew growth to parasitize. In practical terms, treated powdery mildew patches gradually shrink and produce fewer new spores, reducing the disease pressure in the crop.
Importantly, Ampelomyces not only attacks the active, feeding stage of powdery mildew, but can also invade its resting structures. Powdery mildews form sexual spore-bearing structures called chasmothecia (cleistothecia) to survive winter. A. quisqualis has been observed penetrating and developing inside young chasmothecia as well, effectively sterilizing them. By infesting the overwintering stage, Ampelomyces potentially reduces the initial inoculum that causes new infections in the next growing season, a valuable trait for long-term management.
Efficacy: While Ampelomyces works more slowly than chemical fungicides, it can significantly suppress powdery mildew epidemics. In one classic study, greenhouse cucumbers treated with A. quisqualis had much lower mildew coverage compared to untreated plants. Jarvis and Slingsby (1977) reported that periodic applications of Ampelomyces conidial suspensions effectively controlled cucumber powdery mildew in greenhouses, marking the first successful use of a mycoparasite for disease control in a crop. Since then, numerous trials on crops like grapevine, tomato, strawberry, pepper, and rose have shown that A. quisqualis can reduce mildew severity under a range of conditions[1]. Growers have observed that plants treated early (preventatively or at first signs of mildew) benefit the most – the mycoparasite keeps the pathogen in check by curbing its reproduction. The result is often a slower disease spread and lower overall mildew levels, translating to healthier foliage and improved yield or quality.
That said, Ampelomyces is rarely a stand-alone “cure” for heavy mildew infestations. Its best use is preventive or in the very early stages of disease development. Because the fungus needs time to infect and destroy the mildew (often a few days per colony), heavy infections may persist for a while even after Ampelomyces application. In agricultural practice, Ampelomyces is thus used as one component of an integrated strategy (often alternating with low doses of fungicides or used in specific windows of the season – see Section 5 on IPM synergy). Nonetheless, its ability to self-propagate (through pycnidia and secondary infections) is a major advantage – under favorable conditions, one introduction of Ampelomyces can spread and continue suppressing mildew without additional input.In summary, Ampelomyces quisqualis offers a natural and effective means to biologically suppress powdery mildews. By directly attacking the mildew fungi on plants, it provides farmers and gardeners a way to reduce disease with less reliance on chemical sprays, aligning well with sustainable and organic farming principles.
Formulated Biopesticide Products#
The promising biocontrol capabilities of Ampelomyces quisqualis have led to the development of commercial biopesticide products based on this fungus. The most well-known product is marketed under the name AQ10®. AQ10 contains a specific strain of Ampelomyces quisqualis (isolate M-10) that was originally isolated in Israel and deposited in culture collections (e.g., CNCM I-807). It was one of the first microbial fungicides developed for powdery mildew control, receiving registration in the 1990s.
Product formulation: AQ10 is typically formulated as a water-dispersible granule (WG). The active ingredient is A. quisqualis spores at about 58% by weight, with the remainder being carriers and additives to stabilize the product. Each gram of product contains a minimum of ~5×10^9 viable conidia of Ampelomyces. These spores are produced via fermentation and then processed into a dry granular form that can be mixed with water and sprayed. The formulation is designed to keep the spores dormant and stable during storage (often at cool temperatures) and then activate them upon rehydration in the spray tank. Notably, AQ10 contains no live yeast or bacterial additives – it is a pure fungal product, and it is non-genetically-modified (the strain was selected from nature).
Use and registration: AQ10 has been approved in many regions (North America, Europe, and beyond) for use on a variety of crops. For example, in the United States and Canada it can be used on grapes, cucurbits (cucumbers, melons, squash), tomatoes, strawberries, and ornamentals, among others. In the EU and UK, it’s similarly registered for protected (greenhouse) and field crops like cucumbers, tomatoes, eggplant, peppers, and even ornamentals like roses. Because it is a natural organism, AQ10 is exempt from residue tolerances – meaning it does not require a waiting period before harvest (it has a 0-day pre-harvest interval). This makes it very attractive for late-season use or on crops where pesticide residues are a concern (e.g., table grapes or fresh vegetables).
Application of AQ10 is usually recommended at the early stages of powdery mildew infection. The product works best as a preventive or when the first mildew spots are observed, so that Ampelomyces can establish and stop the mildew from spreading. It is typically applied at intervals (e.g., every 7–10 days) for a few cycles. A typical use rate might be around 20–50 grams per hectare, diluted in sufficient water to ensure good coverage of the foliage (since Ampelomyces must contact the mildew colonies). Good coverage is crucial – the spray needs to distribute living spores to the microscopic mildew colonies on the leaves. Once applied, the environmental conditions play a role: high humidity and moderate temperatures (around 20–30°C) are ideal for spore germination and infection. Therefore, applicators often time the sprays for evening or early morning when humidity is highest and sunlight (which can desiccate spores) is low.
Several other Ampelomyces-based products have appeared in different markets, often under different trade names. For instance, a product called “Q-fect” has been used in South Korea, containing a strain of Ampelomyces from a Korean isolate. In India, formulations like “Bio-Dewcon” or “Dr. Bacto’s Ampelo” have been advertised, indicating the global interest in this biocontrol agent. Regardless of the brand, the core of these products is the deployment of Ampelomyces quisqualis spores as a living fungicide against powdery mildew.
Advantages in practice: Ampelomyces-based biofungicides are popular in integrated and organic production systems for several reasons. They leave no chemical residues on the crop, and are safe to non-target organisms – for example, they do not harm beneficial insects or predatory mites, which is important in greenhouses where biological insect controls are used in tandem. They also have no known phytotoxicity on plants when used as directed. However, users must be aware that because Ampelomyces is a living organism, its performance can be inconsistent if conditions aren’t suitable (see Section 6 on challenges). Proper handling (storing cool and dry, using before expiration) and careful application (ensuring humidity, avoiding tank-mixing with incompatible chemicals like certain fungicides – e.g., sulfur or strobilurins can inhibit Ampelomyces) are necessary to get good results.In summary, the commercialization of Ampelomyces quisqualis as AQ10 and similar products has provided farmers with a practical tool for sustainable crop protection. It exemplifies how a naturally occurring antagonist can be harnessed and formulated to manage plant diseases in an environmentally friendly way.
Enzymatic and Biochemical Mechanisms#
The interaction between Ampelomyces quisqualis and powdery mildew fungi is a complex host-parasite relationship that involves both physical invasion and biochemical attack. Unlike some antagonistic fungi (e.g., Trichoderma species) that produce antibiotics or toxins to kill their targets, Ampelomyces is primarily a contact parasite – it must enter the powdery mildew mycelium to exert its effects. Notably, A. quisqualis is not known to produce significant toxins against its host. Earlier research did not detect any toxic metabolites from Ampelomyces that could directly kill mildew fungi in the absence of physical contact. (In contrast, some other fungi parasitic on sclerotia, like Coniothyrium minitans, do produce toxins as part of their mode of action.) This implies that Ampelomyces relies on penetration and enzymatic degradation to parasitize powdery mildews.
Microscopic studies (dating back to the work of de Bary in 1870s and later researchers) show that Ampelomyces hyphae grow inside the cells of the powdery mildew fungus, moving through the septal pores from cell to cell. To achieve this, Ampelomyces likely employs an arsenal of cell wall-degrading enzymes (CWDEs). Fungal cell walls are composed mainly of chitin, β-glucans, proteins, and other polysaccharides. Ampelomyces has been shown to produce enzymes that target these components:
- Chitinases: Enzymes that break down chitin (β-1,4-linked N-acetylglucosamine polymers). Studies detected N-acetylglucosaminidase (a type of chitinase) activity in cultures of A. quisqualis, indicating it can degrade chitin, a major component of fungal cell walls.
- β-1,3-Glucanases: These enzymes cleave β-1,3-glucans, another key component of many fungal walls. A landmark discovery by Rotem et al. (1999) was the identification of an exo-β-1,3-glucanase gene (exgA) in Ampelomyces, which is expressed both in culture and during infection of powdery mildew. Culture filtrates containing this enzyme could even digest the cell walls of powdery mildew hyphae in vitro, underscoring its importance. The β-1,3-glucanase likely helps Ampelomyces perforate the mildew’s cell wall and also to digest it from the inside.
- Proteases: Enzymes that break down proteins. Ampelomyces produces proteolytic enzymes that may degrade structural proteins or enzymes in the host fungus. Protease activity has been recorded during Ampelomyces infections, suggesting a role in breaking down host cytoplasm or disabling host defenses.
- Other enzymes: Researchers have found evidence of β-1,4-glucanase (which could target cellulose-like components or other glucans), β-glucosidase, acid phosphatases, and ribonucleases being produced by Ampelomyces during mycoparasitism. These enzymes could help in digesting the contents of the mildew cells (for example, ribonucleases might break down the host RNA, and acid phosphatases might help scavenge phosphorus from the host).
The combined action of these hydrolytic enzymes effectively turns the powdery mildew’s own cells into a nutrient source for Ampelomyces. In the later stages of infection (5–10 days after Ampelomyces ingress), the mildew’s cytoplasm is observed to be degenerated – a sign that Ampelomyces has consumed or destroyed the host cell contents. At this point the interaction becomes necrotrophic, meaning Ampelomyces is feeding on killed host tissue (initially it is more biotrophic, growing within living host cells without immediately killing them).
Recognition and germination: Before any of the above can happen, Ampelomyces conidia need to recognize the presence of a suitable host. Intriguingly, studies have shown that germination of A. quisqualis spores is stimulated by substances from powdery mildew. For example, a water-soluble compound from powdery mildew conidia can trigger Ampelomyces conidia to germinate in vitro. This suggests a chemical cross-talk: Ampelomyces essentially “smells” its host. This adaptation ensures Ampelomyces spores remain dormant in the absence of a host (which is energy-conserving), and then spring into action when near a powdery mildew colony. Once germinated, the Ampelomyces germ tube grows directionally, apparently chemotropically or via random contact, until it finds the powdery mildew hypha. Often, a small appressorium-like swelling is formed where Ampelomyces attaches to the host wall, indicating a focused point of attack.
Molecular insights: Modern molecular biology has shed more light on Ampelomyces–mildew interactions. A transcriptomic study (Siozios et al. 2015) examined gene expression in A. quisqualis during spore germination in the presence of host (mildew) cues. It revealed that over 1,500 genes changed expression, with many upregulated genes encoding secreted enzymes and putative virulence factors[4]. These included secreted proteases and lipases, as well as proteins related to toxin biosynthesis and other pathogenicity-related processes[4]. The mention of toxin-related genes was interesting – even though Ampelomyces is not known to excrete toxins in culture, it possesses genes that are often found in plant pathogens that do produce toxins. This could mean Ampelomyces has latent or subtle secondary metabolites that aid infection, or that its ancestors had such capabilities (see the Spotlight study in Section 7). Additionally, genes for oxygenases and peptidases were induced, possibly linked to overcoming host oxidative responses or digesting host proteins.
All these data suggest that Ampelomyces quisqualis employs a multifaceted approach: mechanical pressure + enzymatic degradation. By breaking down the structural barriers of its host and then consuming its contents, Ampelomyces effectively neutralizes the powdery mildew fungus. This mode of action is highly specific – it doesn’t harm plant tissue or other organisms, only the mildew fungi – which is ideal for a biocontrol agent. It’s also worth noting that because Ampelomyces attacks multiple components of the mildew (multiple enzymes targeting different polymers), it would be very hard for powdery mildew fungi to develop resistance to Ampelomyces (unlike chemical fungicides that target a single protein or pathway)[1]. This makes A. quisqualis a robust long-term ally in the fight against powdery mildew.
Synergy in Integrated Pest Management#
In modern Integrated Pest Management (IPM) programs, the goal is to combine different control methods to achieve effective pest/disease suppression with minimal chemical use and environmental impact. Ampelomyces quisqualis fits perfectly into IPM for powdery mildew. Rather than replacing chemical fungicides entirely, it often works in synergy with them, and with cultural practices, to keep powdery mildew at bay in a sustainable manner.
Compatibility with fungicides: One might worry that applying a fungicide could kill Ampelomyces, since it’s also a fungus. However, A. quisqualis has shown compatibility with many common fungicides when used carefully. Research has demonstrated that certain systemic fungicides (for example, DMI fungicides like myclobutanil, or some QoI fungicides) do not significantly hinder Ampelomyces from colonizing powdery mildew when used at recommended rates. In practical terms, this means a grower can spray Ampelomyces and a chemical fungicide in an alternating schedule or even tank-mixed (if the products allow) without losing the benefits of the biocontrol. Indeed, field trials have found that combining Ampelomyces with reduced fungicide programs can control disease as well as, or better than, a full chemical program alone. For instance, an trial in Italy alternated AQ10 applications with a standard fungicide on cucumbers, and the integrated approach provided high powdery mildew control – even slightly better than the fungicide-only regime in that case. This indicates a synergistic effect: the chemical quickly knocks back the existing infection and Ampelomyces keeps suppressing any residual or returning mildew, plus potentially slows the development of resistance.
Resistance management: One key advantage of incorporating Ampelomyces into spray schedules is delay of fungicide resistance in the pathogen. Powdery mildew fungi notoriously develop resistance to fungicides (multiple cases of resistance to various fungicide classes have been documented). Because Ampelomyces attacks the mildew in a completely different manner (parasitism rather than a biochemical inhibition of a single target), mildew strains cannot “mutate” to avoid being eaten. By using Ampelomyces, the overall number of chemical sprays can be reduced, or lower doses can be used, which in turn reduces the selective pressure for resistant strains in the mildew population. Some programs use AQ10 in rotations specifically to break up the continuous use of fungicides – for example, two sprays of a fungicide, then one spray of AQ10, then repeat. This integrated approach has been recommended in grape IPM and other crops to maintain effective control over many seasons.
Cultural and biological compatibility: Ampelomyces biocontrol is also compatible with other IPM tactics. Culturally, since it thrives in humid conditions, growers can time irrigation or misting to coincide with Ampelomyces applications (provided it doesn’t excessively promote the mildew itself). In greenhouse settings, ensuring a humid period after application (e.g., by closing vents for a few hours in the evening) can enhance the infection of mildew by Ampelomyces. Conversely, practices like removing heavily mildewed leaves before an Ampelomyces application can help by reducing the densest pathogen load and allowing Ampelomyces to focus on smaller colonies.
When it comes to other biocontrol agents, Ampelomyces is generally friendly. It only attacks powdery mildews, so it won’t interfere with beneficial insects (like ladybugs, parasitic wasps, predatory mites used for insect control) or with bacteria-based biopesticides (like Bacillus subtilis QST strains or Streptomyces products that also target foliar diseases). In fact, Ampelomyces has zero known toxicity to insects or animals. This means it can be used in tandem with biological insect controls – a critical aspect in greenhouse IPM where you often combat both insect pests and diseases simultaneously. For example, one can release predatory mites for spider mite control and spray Ampelomyces for mildew without any harm to the mites.
Application timing and technique: For synergy to be maximized, Ampelomyces must be applied under the right conditions (as touched on earlier). A common recommendation in IPM guides is: apply in the evening or night when relative humidity is rising. This ensures the spores get the moisture they need to germinate on the mildew. If used outdoors, it’s often suggested to apply Ampelomyces on a day when some dew or light rain is expected in the evening, as natural moisture helps spread and activate it (though heavy rain immediately after could wash spores off, so timing is nuanced). In protected environments, humidifying the crop after application (without causing runoff) can improve efficacy. Adjuvants (spreaders/stickers) that are compatible can also be added to improve coverage, especially under less humid conditions. Some studies tested special adjuvants (e.g., vegetable oils or polymer stickers) with Ampelomyces; while these didn’t increase the infection rate dramatically, they did not harm the spores either. In low-humidity climates, an adjuvant can help Ampelomyces by slowing desiccation of spores on the leaf.
IPM outcomes: When Ampelomyces is well-integrated into an IPM program, growers have reported:
- Reduced number of fungicide applications needed to keep powdery mildew below damaging levels.
- Maintenance of efficacy where purely chemical programs were failing due to resistant mildew strains.
- Improved yields or quality in sensitive crops (e.g., less russeting on grapes or strawberries, since fewer sulfur or harsh fungicide sprays are used).
- Compliance with organic certification or residue regulations when Ampelomyces is used in place of certain chemicals.
As an example, an IPM program in greenhouse strawberries might involve: planting resistant or tolerant varieties (cultural control), monitoring humidity and venting to avoid extreme conducive conditions (environmental control), releasing Ampelomyces via a biopesticide like AQ10 at first detection of mildew (biocontrol), and using one or two targeted fungicide sprays during peak pressure (chemical control). This multi-pronged approach keeps the disease manageable and is sustainable long-term.
In conclusion, Ampelomyces quisqualis exemplifies the synergy of biological and chemical controls – it works best not in isolation, but as a team player in IPM. Its addition to disease management programs helps in rationalizing fungicide use, delaying resistance, and achieving adequate control with a softer environmental footprint. Growers adopting IPM have found that embracing such biocontrol agents is an investment in the future – as regulatory pressures and resistance issues limit fungicides, having a reliable bio-tool like Ampelomyces is invaluable for sustainable agriculture.
Challenges and Future Potential#
While Ampelomyces quisqualis is a powerful tool for managing powdery mildews, there are several challenges associated with its use and biology. At the same time, ongoing research is addressing these issues, and there is significant future potential to enhance the effectiveness of this mycoparasite.
Variable Efficacy: One of the main challenges in using Ampelomyces as a biocontrol agent is its sometimes inconsistent performance in the field. Unlike a chemical fungicide that can be broadly reliable (weather permitting), Ampelomyces is a living organism whose success can be influenced by many factors. Field trials have reported mixed results: in some cases A. quisqualis provided excellent control of powdery mildew, equivalent to 50–70% disease reduction or more, while in other trials the disease suppression was marginal. These discrepancies can arise from differences in climate, crop, mildew species, application technique, and even the Ampelomyces strain used. For instance, an experiment might show good control on grapevine in a cool, humid region, but only modest effects on zucchini squash in a hot, dry climate. Such variability means growers must learn the best way to use Ampelomyces in their specific conditions – it may not be a one-size-fits-all solution.
Environmental Dependence: Ampelomyces’ activity is strongly influenced by environmental conditions, especially moisture. High relative humidity (ideally above ~80%) for several hours after application is needed for optimal germination and infection. If conditions are too dry, the spores may not germinate at all, or germinate and die before finding the host. Similarly, heavy rain can wash the spores off leaves, and extreme heat or UV exposure can inactivate them. Therefore, the window for effective application can be narrow. In outdoor crops, weather unpredictability adds an element of risk – a well-timed Ampelomyces spray could be rendered ineffective by an unexpected dry wind or scorching sun. Some growers mitigate this by applying in the evening (to take advantage of overnight dew) or using mist irrigation after application. The reliance on specific microclimatic conditions is a hurdle to consistency. However, this is being addressed by formulation improvements (e.g., encapsulating spores or adding protective carriers to keep them moist longer) and by better forecasting tools that advise when conditions will favor Ampelomyces infection.
Slow Action and Coverage: Ampelomyces is a relatively slow-growing fungus, especially compared to the fast-sporulating powdery mildews. On artificial media, A. quisqualis colonies expand only ~0.5–1 mm per day for the slow-growing strains (those considered A. quisqualis sensu stricto). This slow growth is reflected in its biocontrol action – it takes several days to completely parasitize a mildew colony. During this time, the powdery mildew may continue to cause some damage or even spread slightly. Thus, Ampelomyces is not a “knockdown” control; it’s more of a suppressive agent that needs to be present continuously. Achieving thorough coverage of a crop with Ampelomyces is crucial so that it can colonize all or most mildew colonies and gradually reduce the epidemic. If some pockets of mildew remain untreated (for example, in dense canopies where spray coverage is poor), those can continue to thrive and undo the benefits. This challenge is common to many biocontrols – they lack the curative punch of chemicals but excel in preventive suppression. Farmers must be educated that Ampelomyces will reduce sporulation and spread of mildew, but existing severe infections won’t disappear overnight. In practice, this might mean integrating a low-rate fungicide to burn back heavy infestations, then relying on Ampelomyces for ongoing control.
Strain and Genetic Diversity: As highlighted earlier, what we call Ampelomyces quisqualis is actually a genetically diverse group of strains. Molecular studies using DNA sequencing have revealed at least 4–7 distinct lineages (clades) within what was historically identified as A. quisqualis. These lineages might correspond to different species – indeed, the genus Ampelomyces likely contains multiple species that are very similar morphologically. This has several implications:
- Host Specialization: Some Ampelomyces lineages may be better adapted to certain powdery mildew hosts. For example, a strain isolated from grape powdery mildew might infect grape mildew very effectively, but perhaps not mildew on cucurbits as aggressively (and vice versa). There is evidence that while Ampelomyces strains can infect many hosts, they often show a degree of preference or superior performance on their original host type. This specialization is not absolute (no strict one-strain-one-host rule), but it suggests that “matching” strain to crop could improve results. Currently, the commercial strain AQ10 (isolate M-10) is used on all crops, but in the future, we might see crop-specific Ampelomyces strains being developed.
- Variability in Virulence: Not all Ampelomyces strains are equally virulent or aggressive. Researchers like D. Angeli and colleagues found that different strains produced different levels of hydrolytic enzymes and had varying biocontrol efficacy in tests. This means there is an opportunity to select or breed superior strains. The strain in AQ10 was chosen in part for its ability to grow and sporulate well in fermentation, but perhaps an even more virulent strain exists in nature that could provide stronger biocontrol if it can be mass-produced. Ongoing exploration in different regions (e.g., recent studies in Japan isolated many Ampelomyces strains from local mildews and assessed their mycoparasitic activity) will likely enrich the pool of candidates for new biocontrol products.
- Taxonomy and Regulation: The unresolved taxonomy could also pose a regulatory challenge. If, say, AQ10’s strain is reclassified as Ampelomyces quisqualis “Clade 1” and another product uses a strain from Clade 4 (which might even deserve a different species name), regulators might treat them as different active ingredients. A clear taxonomic revision (which scientists are calling for) will help streamline how we refer to and regulate these biocontrol agents.
Mass Production and Shelf Life: Producing Ampelomyces spores on an industrial scale and formulating them into a stable product has been a challenge, though much progress has been made. A. quisqualis is traditionally grown on solid substrates (like grain or agar), but newer methods involve liquid fermentation. Research has managed to produce A. quisqualis conidia in submerged culture (liquid broth), which is not trivial because this fungus naturally sporulates on or in a host, not freely in liquid. Innovations such as optimized nutrient media and subtle stresses have been used to induce sporulation in fermentation tanks. Even so, achieving high spore concentrations and harvesting them without damage requires specialized techniques. Once harvested, the spores need to be formulated with carriers that keep them alive. Desiccation can kill spores, so formulating as a slightly moist granule or with water-absorbing protectants is common. Companies have worked on improving shelf-life – for instance, Angeli et al. (2017) reported formulation improvements that extended Ampelomyces conidia viability significantly. Currently, most products have a shelf life of 1–2 years under cool storage. Continuing improvements in formulation (microencapsulation, oil dispersion formulations, etc.) are a future avenue to make Ampelomyces products even more user-friendly and reliable.
Future Potential – Research and Innovation: The future of Ampelomyces in crop protection looks bright thanks to scientific advancements:
- Genomics and Bioengineering: The genome of A. quisqualis has now been sequenced (see Spotlight on Research in Section 7), revealing genes and pathways that were previously unknown[5]. This opens up possibilities to identify genetic markers for hypervirulence or environmental tolerance. In the long run, scientists could potentially genetically improve Ampelomyces strains – for example, by knocking out a gene that limits its growth or by enhancing a gene for chitinase production, to create a strain that kills mildew even faster. Of course, GMO biocontrol agents would have regulatory hurdles, so an alternative is using the genomic knowledge to guide classical strain selection (screening natural variants for those with desired traits).
- Formulation Technology: Future formulations might include dry spores in capsules that protect them from UV and release them upon wetting, or combination products where Ampelomyces is mixed with a helper organism (for example, a yeast that creates a humid microenvironment on the leaf or a nutrient that boosts Ampelomyces germination).
- New Application Methods: Researchers are also looking at novel ways to apply Ampelomyces. One idea is endophytic delivery – finding endophytic fungi or bacteria that could carry Ampelomyces into plants or at least onto leaf surfaces in a more natural way. Another concept is formulating Ampelomyces in slow-release devices or as part of a consortium spray (mixing it with other beneficial microbes that together target multiple diseases or pests).
- Broader Target Range: While Ampelomyces is specific to powdery mildews, the concept of mycoparasites in IPM encourages looking for analogous solutions for other diseases. Ampelomyces itself might not control, say, downy mildews or rusts (unrelated groups of pathogens), but the successful use of Ampelomyces is a proof-of-concept that could drive development of hyperparasites for those diseases. In the realm of powdery mildews, Ampelomyces will likely remain the flagship hyperparasite, but one could imagine combining it with other antagonists (like certain beneficial microbes that live on the leaf surface and inhibit mildew germination) for a multi-modal attack on the pathogen.
Sustainable agriculture significance: Ultimately, the continued refinement of Ampelomyces quisqualis for crop protection aligns with the global push for sustainability. It reduces growers’ dependency on synthetic chemicals, cuts down residues in food and environment, and exploits a naturally occurring ecological interaction (parasite on parasite) to our advantage. Challenges like environmental dependence and slow action are being mitigated by smarter use (predictive models to time applications, etc.) and by integrating Ampelomyces with other measures so its weaknesses are covered by others’ strengths.
In conclusion, Ampelomyces has proven its value over decades, and with ongoing research, its future potential is even greater. We can expect more efficient strains, more robust products, and deeper understanding to emerge, all of which will cement Ampelomyces quisqualis as a cornerstone of eco-friendly disease management in agriculture.
Spotlight on Research:#
Brief Overview#
One particular study that highlights the strides being made in Ampelomyces research is the first whole-genome sequencing of Ampelomyces quisqualis, published by Huth et al. in 2021. Until recently, the genetic and evolutionary understanding of A. quisqualis was limited – scientists knew it was effective as a biocontrol and had documented its enzyme production and diversity, but its entire genome had not been decoded. Huth et al. tackled this by sequencing a high-quality draft genome of an A. quisqualis strain (designated BRIP 72107) isolated in Queensland, Australia. This was a breakthrough because it provided a comprehensive look at the fungus’s genetic blueprint. Their research had twin aims: (1) to compare the genome of Ampelomyces to other fungi (especially to see if it has traits in common with plant pathogens or other lifestyles), and (2) to clarify the taxonomy and relationships within the Ampelomyces genus by using genome data.
The study was notable for applying cutting-edge sequencing technologies (a combination of long-read Nanopore sequencing and short-read Illumina sequencing) to assemble the genome into relatively few pieces (scaffolds) with high completeness[5]. They also conducted phylogenetic analyses using not just the traditional ITS region but also genome-wide data to see how this strain relates to others. One interesting aspect is that they compared their genome (from an Australian strain) to a previously released genome sequence of a different Ampelomyces strain from elsewhere – giving a genetic basis to the concept of different lineages within Ampelomyces. In essence, this study represented a major step from classical microbiology into the genomic era for A. quisqualis.
Key Insights#
The genome study by Huth et al. yielded several key insights:
- Genome Size and Structure: The genome of Ampelomyces quisqualis was found to be approximately 40.4 million base-pairs (Mb) in size, organized into 24 major scaffolds (chromosome-like fragments)[5]. This size is moderate for a fungus and provides an inventory of thousands of genes. A striking discovery was the “bipartite” (two-part) structure of the genome. Rather than being uniform, the Ampelomyces genome is mosaic, with regions of typical GC content interspersed with regions that are AT-rich and gene-poor. Specifically, large stretches of AT-rich DNA (some up to 100–200 kb long) were identified, comprising a significant portion of the genome. Such AT-rich, repeat-rich regions are reminiscent of the so-called “two-speed genome” found in many plant pathogenic fungi (for example, in Leptosphaeria maculans, a rapeseed pathogen, and others). In those pathogens, AT-rich regions often harbor transposable elements and effector genes and are thought to allow rapid evolution. Finding this in Ampelomyces suggests it may share an evolutionary strategy with pathogenic fungi – possibly reflecting an ancestor that was a plant pathogen or at least that Ampelomyces has the genomic toolkit to rapidly adapt, perhaps to different hosts.
- Evolutionary Clues: The presence of a bipartite genome and other genomic features led the authors to propose that Ampelomyces evolved from plant-pathogenic ancestors. This is a fascinating hypothesis: perhaps at some point in evolution, a fungus that used to infect plants switched to infecting the fungi on those plants. The genome supports this idea by the similarity of its structure to plant pathogens and possibly by containing relics of pathogenicity genes. Huth et al. noted that this could explain why Ampelomyces can reside inside other fungi – it may have repurposed a skill set that once let it live in plant tissue.
- Multiple Species (MOTUs): By comparing genomic (and ITS) data of different Ampelomyces strains, Huth et al. reinforced that Ampelomyces is not one single homogeneous species. In their analysis, the Australian strain BRIP 72107 belonged to a clade (which they call MOTU1) that includes the commercial AQ10 strain. However, another sequenced strain (from a study by Haridas et al. 2020, labeled HMLAC 05119) fell into a different clade (MOTU4), and even another strain used in Korea (strain 94013, product Q-fect) was also in MOTU4. This means AQ10’s strain and the Australian strain are essentially the same species (conspecific), while those other strains represent at least one other species. In total, they identified nine well-supported clades (MOTUs) among Ampelomyces isolates analyzed worldwide. This comprehensive phylogeny underscores the need to revise Ampelomyces taxonomy – we may eventually have multiple Ampelomyces species names instead of calling all of them A. quisqualis. For biocontrol, this is key information: it tells us that some strains (like AQ10’s) are genetically distinct from others (like the Korean one), which might explain differences in performance or host range that have been observed.
- Gene Repertoire: Although the study’s main focus was on genome structure and phylogeny, having the genome allowed the authors to annotate thousands of genes. They reported that A. quisqualis has all the expected genes for a filamentous fungus, and importantly, it carries a range of genes that could be tied to its parasitic lifestyle – for example, genes for cell wall degrading enzymes, stress tolerance, and secondary metabolism (though the latter seem less expressed, aligning with the fact it’s not known to produce toxins vigorously)[4]. One particularly interesting find was that Ampelomyces lacks certain genes that many free-living fungi have, possibly due to its specialized niche living inside other fungi. Such genomic insights can direct future studies to specific gene families (like chitinases or effectors) to see how Ampelomyces attacks its host.
Why This Matters#
This genome sequencing study is significant for both fundamental and applied reasons:
- Improving Biocontrol: Understanding the genome of Ampelomyces can directly contribute to making it a better biocontrol agent. For instance, if we know all the enzymes it uses to kill powdery mildew, we might be able to screen natural populations for strains that overproduce a particular enzyme, thereby killing the host faster. Alternatively, if we discover a gene that limits spore production, we could target that in strain improvement programs. In the shorter term, the genome allows us to develop molecular markers – tools to quickly identify different strains or lineages of Ampelomyces. This is useful if a farmer or researcher wants to monitor how well an introduced strain (like AQ10) is establishing in the field, or if native strains are present. It also helps in quality control of commercial products: DNA tests can confirm that the product contains the correct strain and is free of contaminants.
- Evolutionary Insight: From a scientific standpoint, the hypothesis that Ampelomyces may have evolved from plant pathogens blurs the line we traditionally draw between “good” fungi and “bad” fungi in agriculture. It reminds us that evolution can be opportunistic – a plant pathogen might lose the ability to attack the plant but gain the ability to parasitize a fellow fungus. This insight broadens our understanding of fungal ecology and could guide searches for other biocontrol agents. If hyperparasites like Ampelomyces originate from plant pathogens, perhaps we should look among known plant pathogens for candidates that could switch to attacking other pathogens. It’s a bit of a mind-bending concept: the enemy of our enemy (a biocontrol fungus) might itself be a former enemy of plants. The genome data provide evidence for this trajectory by showing shared genome characteristics with plant-pathogenic relatives.
- Taxonomy and Regulatory Clarity: Practically, knowing there are multiple clades (MOTUs) of Ampelomyces helps avoid confusion. For example, if a new biocontrol product is developed from a different Ampelomyces strain, one can use genomic data to clearly differentiate it from AQ10’s strain so that both can be used optimally (and regulatory agencies can register them properly). It can also prevent misattribution if a research result or field observation is tied to one strain but not another. In the long run, as taxonomy is revised, publications and labels will likely start specifying the lineage (perhaps A. quisqualis “Clade 1” vs Ampelomyces sp. Clade 4, until formal names are given).
- Foundation for Future Research: This study provides a foundation for a lot of follow-up research. For instance, researchers can now perform comparative genomics: comparing Ampelomyces’ genome to that of the powdery mildew fungi it attacks. What genes does a parasite have that its host doesn’t, and vice versa? Is there a coevolution signal (e.g., mildew genomes adapting to resist hyperparasites)? Also, transcriptomic and proteomic studies can now map onto the genome. The 2015 transcriptome study (Siozios et al.) found many upregulated genes[4] – now scientists can pinpoint their exact location and context in the genome, and perhaps identify their regulatory elements or whether they reside in AT-rich regions (which might mean they are fast-evolving genes).
- Public Perception and Acceptance: Although not a direct scientific outcome, having detailed genomic information on a biocontrol agent can sometimes help with public acceptance and regulatory approval. It shows that the organism is being well-studied and monitored at a molecular level, which can alleviate concerns about “What exactly are we spraying in the environment?”. It also demonstrates that biocontrols are being developed with as much rigor as pharmaceuticals, which can boost confidence in their use.
In sum, the Huth et al. (2021) genome study matters because it propels Ampelomyces quisqualis into the genomic age, equipping researchers and industry with knowledge to refine and exploit this fungus even more effectively. It serves as a model for how to dissect a biocontrol agent’s biology and how to link fundamental evolutionary insights to practical improvements in agriculture.
Summary Table: Spotlight Study#
| Category | Details |
| Lead Researchers | Lauren Huth; Gavin J. Ash; Alexander Idnurm; Levente Kiss; Niloofar Vaghefi – a team of plant pathology and mycology researchers. |
| Affiliations | Centre for Crop Health, University of Southern Queensland, Australia; School of BioSciences, University of Melbourne, Australia (among others). |
| Research Focus | Sequencing and analyzing the genome of Ampelomyces quisqualis to understand its genetic make-up, evolutionary origin, and the genomic basis of its mycoparasitic lifestyle. |
| Key Breakthroughs | – Produced the first high-quality genome assembly of A. quisqualis (≈40.38 Mb, 24 scaffolds, 96% complete)[5]. – Discovered a “bipartite” genome structure: gene-rich, GC-normal regions interspersed with large AT-rich, repeat-laden regions. Such a two-speed genome architecture is a hallmark of many plant pathogenic fungi. – Found genomic evidence suggesting Ampelomyces evolved from plant-pathogenic ancestors (shared genome features with pathogens). – Clarified that Ampelomyces consists of multiple phylogenetic lineages (at least 9 MOTUs); the sequenced strain belongs to the same clade as the commercial AQ10 strain, whereas another sequenced strain (“HMLAC 05119”) belongs to a different clade. |
| Collaborative Efforts | Involved collaboration between universities in Australia and experts in fungal genomics and pathology. The team combined expertise in biocontrol (Ash, Kiss) and molecular genetics (Idnurm, Vaghefi) to interpret the data, and referenced work by international colleagues (e.g., in Hungary, Japan) for comparative analysis. |
| Published Work | Published in Genome Biology and Evolution (Oxford University Press) in August 2021. Full reference: Huth et al., 2021, Genome Biol. Evol. 13(8): evab182. This is a peer-reviewed, high-impact journal focusing on genomic and evolutionary biology. |
| Perspective | This study provides a new perspective on Ampelomyces: rather than a simple biocontrol agent, it is a fungus with a complex genome that hints at a pathogenic past. It bridges our understanding between plant pathogens and hyperparasites, suggesting evolutionary fluidity between these roles. For biocontrol practitioners, it legitimizes Ampelomyces as a thoroughly studied organism (genetically) and paves the way for genome-informed improvements in its use. |
| Publication Date | Online on August 3, 2021 (print issue August 2021). This makes it a very recent study (as of 2021) offering up-to-date insights. |
| Location | The Ampelomyces strain sequenced (BRIP 72107) was isolated from a powdery mildew on Cestrum (ornamental plant) in Toowoomba, Queensland, Australia. The genome work was carried out in labs at University of Southern Queensland and University of Melbourne (Australia), with the strain stored in the Queensland Plant Pathology Herbarium (BRIP). The study thus had an Australian nexus, though it incorporated comparative data from strains isolated in other countries (Europe, Asia) for the phylogenetic analysis. |
| Key Findings | – A. quisqualis genome ~40.4 Mb, with a bimodal GC content (AT-rich vs GC-rich regions). About ~45–50% of the genome is repetitive/AT-rich, suggesting active transposons and a “two-speed” genome strategy. – Genomic layout and composition are similar to certain plant pathogens (e.g., Leptosphaeria maculans), supporting an evolutionary link. – Phylogenetics: The sequenced strain and the AQ10 biocontrol strain are in the same clade (MOTU1), confirming AQ10’s taxonomic placement. Another commercial strain (“Q-fect” from Korea) and a previously sequenced strain form a separate clade (MOTU4), showing cryptic speciation within Ampelomyces. – Highlighted the need for taxonomic revision: formally, all these strains are called A. quisqualis, but genetically they may be distinct species. A comprehensive polyphasic taxonomy study was suggested. – Provided a catalog of genes: identified genes for cell wall degrading enzymes, stress response, and other factors potentially important for mycoparasitism. This offers targets for future functional studies or strain enhancement. |
Conclusion#
Ampelomyces quisqualis stands out as a success story in the realm of biological control and sustainable agriculture. From its early discovery parasitizing grapevine mildew in the 1800s, to Yarwood’s first biocontrol experiment in 1939, to modern-day formulated products and genomic revelations, A. quisqualis has come a long way. It embodies the principle of “the enemy of my enemy is my friend” – by preying on powdery mildew fungi, this humble fungus has become a valuable friend to farmers and gardeners seeking eco-friendly disease management.
In practical use, Ampelomyces offers a natural, residue-free means to suppress a disease that would otherwise require frequent chemical fungicides. Its role is not to entirely replace chemicals in all situations, but to augment and reduce our reliance on them. By integrating A. quisqualis into IPM programs (timing its application to humidity, rotating it with compatible fungicides, etc.), growers can achieve control of powdery mildew with fewer chemical inputs, thereby mitigating issues like fungicide resistance and environmental contamination[1]. Importantly, Ampelomyces poses no risk to the crop or the consumer – it only targets the pathogenic fungus. This makes it especially appealing for high-value crops (e.g., grapes for wine, where residue could affect fermentation, or vegetables in greenhouse, where worker re-entry and residue are concerns) and for organic farming where synthetic fungicides are prohibited.
From a scientific perspective, A. quisqualis has taught us much about mycoparasitism – the intriguing interactions where fungi fight fungi. Studies on its life cycle revealed how intricately it times its attack (even using chemical signals from the host to germinate), and how it can shut down the reproduction of a pathogen by colonizing its fruiting structures. It has also underscored the complexity hidden in what was once thought to be a single species; the genetic diversity within Ampelomyces is an active area of research that will refine how we deploy these organisms.
Looking forward, the future potential of Ampelomyces quisqualis is promising. With genome-driven insights, we may see next-generation biocontrol products: perhaps strain “cocktails” covering multiple clades to ensure efficacy on all powdery mildew types, or strains bred for specific climates (one more heat-tolerant for tropical use, another more cold-tolerant for use in early spring in temperate regions). There is also interest in exploring Ampelomyces in combination with digital agriculture tools – for example, disease prediction models could trigger an alert to apply Ampelomyces before a powdery mildew outbreak explodes, maximizing its preventive power[1].In conclusion, Ampelomyces quisqualis exemplifies how leveraging natural antagonisms can make crop protection more sustainable. It demonstrates that effective disease control doesn’t always require synthetic chemicals; sometimes it can be achieved by carefully introducing the right microbe at the right time. As research continues and application techniques improve, Ampelomyces is poised to remain a cornerstone of powdery mildew management. It’s a testament to the ingenuity of both nature and science – nature, for evolving a fungus that preys on other fungi, and science, for recognizing and harnessing this trait for the benefit of agriculture. With growing global emphasis on reducing chemical inputs and preserving environmental health, Ampelomyces quisqualis truly is a fungus whose time has come, offering a path to sustainable crop protection for years to come.
References#
- Németh, M.Z.; Seress, D.; Nonomura, T. (2023). Fungi Parasitizing Powdery Mildew Fungi: Ampelomyces Strains as Biocontrol Agents against Powdery Mildews. Agronomy, 13(8): 1991. DOI: 10.3390/agronomy13081991.
- Angeli, D.; Puopolo, G.; Maurhofer, M.; Gessler, C.; Pertot, I. (2012). Is the mycoparasitic activity of Ampelomyces quisqualis biocontrol strains related to phylogeny and hydrolytic enzyme production? Biological Control, 63(3): 348–358.
- Rotem, Y.; Yarden, O.; Barash, I.; Sztejnberg, A. (1999). The mycoparasite Ampelomyces quisqualis expresses exgA encoding an exo-β-1,3-glucanase in culture and during mycoparasitism. Phytopathology, 89(8): 631–638.
- Siozios, S.; Tosi, L.; Ferrarini, A.; Ferrari, A.; Tononi, P.; Bellin, D.; Maurhofer, M.; Gessler, C.; Delledonne, M.; Pertot, I. (2015). Transcriptional reprogramming of the mycoparasitic fungus Ampelomyces quisqualis during the powdery mildew host-induced germination. Phytopathology, 105(2): 199–209pubmed.ncbi.nlm.nih.govpubmed.ncbi.nlm.nih.gov.
- Huth, L.; Ash, G.J.; Idnurm, A.; Kiss, L.; Vaghefi, N. (2021). The “bipartite” structure of the first genome of Ampelomyces quisqualis, a common hyperparasite and biocontrol agent of powdery mildews, may point to its evolutionary origin from plant pathogenic fungi. Genome Biology and Evolution, 13(8): evab182pmc.ncbi.nlm.nih.gov.
- Jarvis, W.R.; Slingsby, K. (1977). The control of powdery mildew of greenhouse cucumber by water sprays and Ampelomyces quisqualis. Plant Disease Reporter, 61: 728–730.
- Sundheim, L. (1982). Control of cucumber powdery mildew by the hyperparasite Ampelomyces quisqualis and fungicides. Plant Pathology, 31: 209–214.
- U.S. Environmental Protection Agency (EPA). (1999). Pesticide Product Label: AQ-10 Biofungicide (Ampelomyces quisqualis isolate M-10). EPA Registration No. 55638-16.
- LS Systems Ltd. (2012). AQ10 Technical Notes (Product Information Bulletin, Intrachem/Biogard), Issue PM-06/12, pp. 1–8.