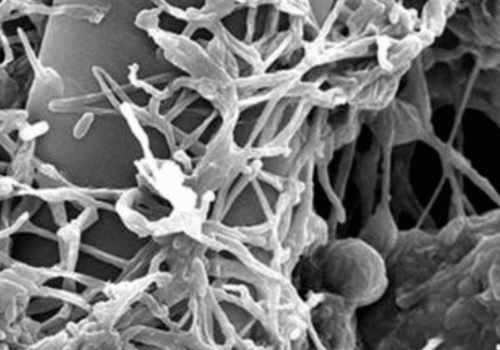
Pochonia chlamydosporia: A Nematode-Parasitic Fungus for Eco-Friendly Crop Protection
Overview of the Microbe Pochonia chlamydosporia is an ascomycete fungus (Hypocreales: Clavicipitaceae) formerly known as Verticillium chlamydosporium[1]. First identified in the 1970s as a natural...
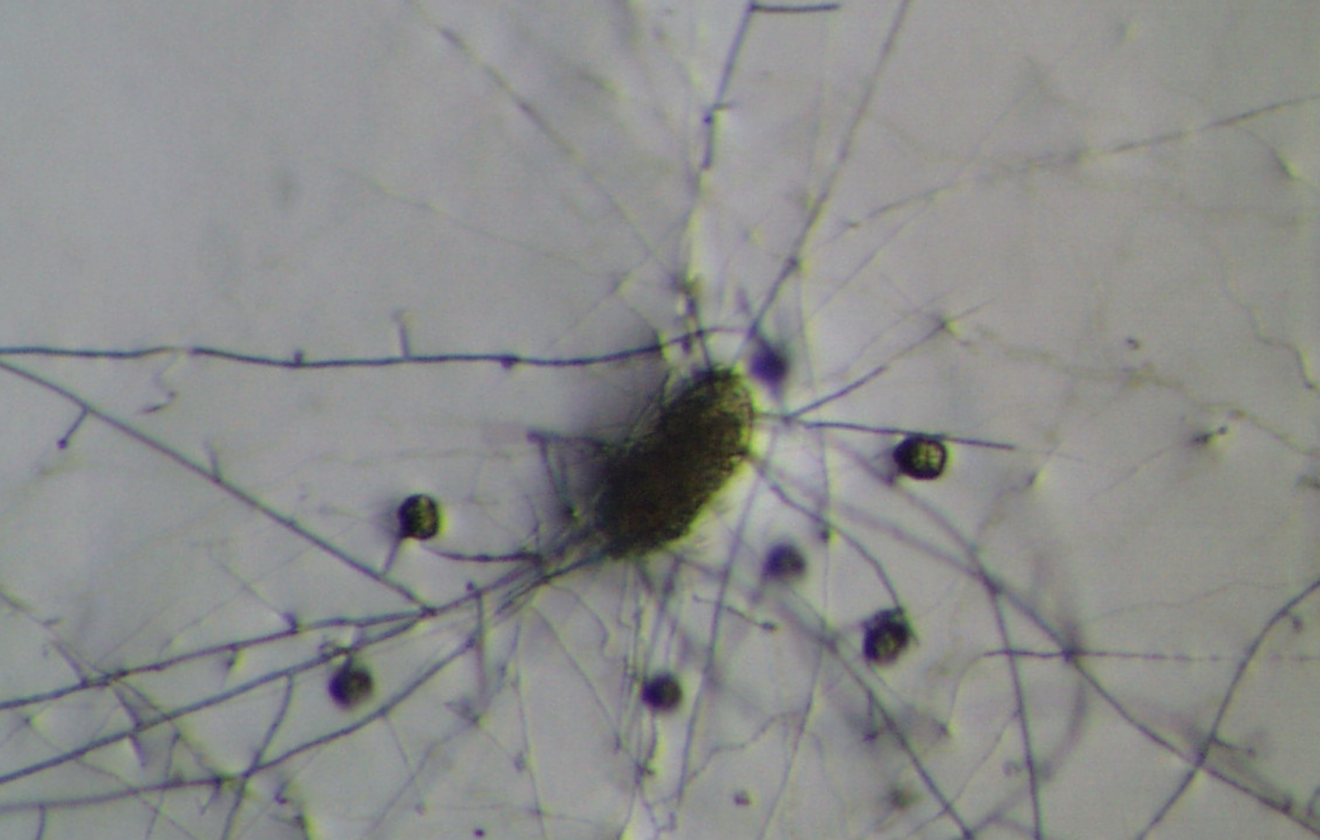
- Overview of the Microbe
- Biocontrol of Plant-Parasitic Nematodes
- Mycoparasitism Mechanisms and Enzymatic Arsenal
- Induced Systemic Resistance and Plant Growth Promotion
- Formulation and Field Application Strategies
- Challenges and Future Outlook
- Spotlight on Research: Pochonia chlamydosporia – Lin et al. (2018)
- Conclusion
- References
- 1
Effect of plant-parasitic nematodes on root growth over time
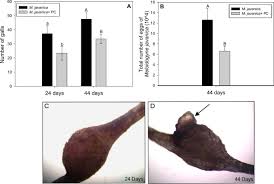
The bar graphs and images show how nematode infection reduces the number and size of plant galls (swollen root areas) over 24 and 44 days. Infected roots show visible damage and fewer galls, especially with certain treatments.
- 2
How plants make key defense and growth compounds
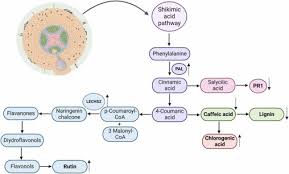
This diagram shows the plant’s internal pathway (starting from the shikimic acid route) for producing important substances like flavonoids, salicylic acid, chlorogenic acid, and lignin.
Overview Table of Pochonia chlamydosporia
- Feature
Description
- Scientific Name
Pochonia chlamydosporia
- Classification
Ascomycete fungus; Phylum Ascomycota; Order Hypocreales
- Habitat
Soil, rhizosphere of diverse crops
- Key Functions
Parasitism of nematode eggs and cysts; organic matter decomposition
- Notable Abilities
Production of chlamydospores; secretion of proteases, chitinases; rhizosphere colonisation
- Applications
Biocontrol of root-knot and cyst nematodes; integrated pest management; soil health enhancement
- Genetic Engineering Potential
Targeted overexpression of lytic enzyme genes; transformation for enhanced rhizosphere fitness
- Challenges
Variable field efficacy; formulation stability; regulatory hurdles
- Future Prospects
Synthetic community design; genomic-assisted strain selection; integration into circular bioeconomy
Overview of the Microbe#
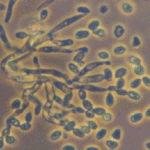
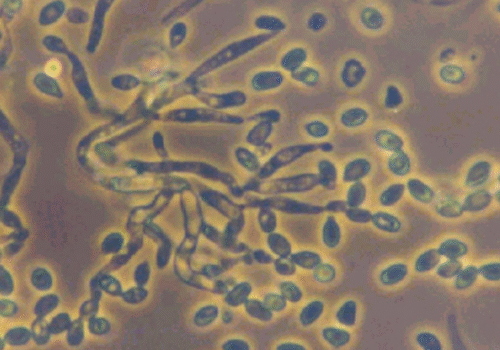
Pochonia chlamydosporia is an ascomycete fungus (Hypocreales: Clavicipitaceae) formerly known as Verticillium chlamydosporium[1]. First identified in the 1970s as a natural parasite of nematode eggs[1], it has since been found worldwide, often in nematode-suppressive soils where it parasitizes root-knot and cyst nematode eggs[1]. The fungus is multitrophic: it can survive saprophytically on organic matter, colonize plant roots as an endophyte, and parasitize nematodes[1][2]. In culture it readily produces hardy chlamydospores (thick-walled spores), which give the species its name and help it persist in soil. P. chlamydosporia is known to colonize the rhizosphere and root tissues of crops (especially cereals and solanaceous plants), where it may even enhance plant defenses[1]. It is a facultative parasite of nematode and some mollusk eggs, and can hyperparasitize other soil fungi[1].
Taxonomically, P. chlamydosporia was described by Goddard and later placed in Pochonia by Zare & Gams (2001)[1]. Several varieties exist (e.g. var. catenulata) that differ in colony morphology and genetic markers. Genomic studies have now assembled its chromosomes, revealing a genome of ~44 Mb with abundant genes for secreted enzymes[9]. These secretome genes suggest a strong adaptation to chitin-rich environments and nematode parasitism. In culture and field surveys, diverse isolates have been recovered from soils worldwide, but only a few strains have been developed into commercial biocontrol products[1].

Biocontrol of Plant-Parasitic Nematodes#
P. chlamydosporia is one of the most-studied biological control agents for plant-parasitic nematodes (PPN)[1]. It targets sedentary endo-parasitic nematodes (which spend most of their life inside roots) by destroying their eggs. Major host genera include root-knot nematodes (Meloidogyne spp.), cyst nematodes (Globodera, Heterodera spp.), and more recently vine and reniform nematodes such as Nacobbus and Rotylenchulus[1]. In greenhouse and field trials, P. chlamydosporia inoculations have consistently reduced nematode reproduction and root damage. For example, inoculation of tomato with P. chlamydosporia (strain PC-170) cut Meloidogyne incognita egg counts and root galls by roughly half compared to controls[4]. In one study, three strains isolated from a suppressive soil all significantly lowered egg masses of M. incognita, with the best isolate reducing eggs by ~50% and also decreasing second-stage juveniles and galling[4].
Field trials have likewise shown promise. A commercial formulation (“POCHAR”) containing P. chlamydosporia applied via irrigation significantly controlled root-knot nematodes on potato and promoted tomato growth under field conditions[3]. In that study, treated plots had markedly fewer nematode-infested tubers and higher plant biomass than untreated plots, demonstrating that P. chlamydosporia can function as an organic alternative to chemical nematicides[3]. Other studies report similar trends in cucumber, lettuce, and other crops. In general, P. chlamydosporia treatments (often used as part of integrated pest management) reduce nematode egg hatch and infection rates, leading to healthier roots and higher yields.
However, biocontrol efficacy can vary with strain and context. Different isolates of P. chlamydosporia show preferences for different nematode hosts, reflecting genetic diversity[7][1]. Some highly virulent strains are favored for specific crops. In practice, P. chlamydosporia is usually applied before planting or in combination with other beneficial microbes (e.g. nematophagous Arthrobotrys or mycorrhizal fungi) to maximize soil colonization[3].
Mycoparasitism Mechanisms and Enzymatic Arsenal#
The biocontrol action of P. chlamydosporia hinges on its ability to parasitize nematode eggs and juveniles. Microscopic studies show that fungal hyphae attach to the surface of eggs/cysts, then penetrate and digest them. A suite of hydrolytic enzymes is secreted to breach the tough, chitin-rich egg shells. Key enzymes include proteases (notably a subtilisin-like serine protease called VCP1) and chitin-degrading enzymes such as chitinases, chitin deacetylases, and chitosanases[5][2]. VCP1 is produced very early in infection, where it degrades the proteinaceous vitelline membrane of nematode eggs[5]. Genetic analyses have revealed that VCP1 from different P. chlamydosporia isolates contains amino acid variations that correlate with the nematode host species, suggesting adaptation of the protease to different egg compositions[7].
Metabolically, P. chlamydosporia is especially adept at chitin and chitosan breakdown[2]. Genomic studies show a large array of secreted chitinases, chitin deacetylases, and chitosanases in its genome[2]. The fungus can grow on chitin substrates and rapidly degrade chitosan, consistent with observations that chitosan is a major component of nematode eggshells[2]. Transcriptomic analyses confirm that during egg infection the fungus upregulates genes for these enzymes. For example, one study found that the presence of nematode eggs triggers increased VCP1 expression in P. chlamydosporia, indicating egg-specific induction of protease secretion[5]. Another found that chitosan (a chitin derivative) strongly modulates fungal gene expression: genes encoding proteases, chitosanases and lectin-like adhesins are upregulated in the presence of chitosan and eggs[8].
Taken together, P. chlamydosporia uses a “cut-and-digest” strategy: fungal hyphae adhere to the egg surface (often mediated by secreted lectin/adhesive proteins), then secrete a cocktail of lytic enzymes that soften and hydrolyze the shell and vitelline layers[8][5]. Genomic analysis identified ~1,750 secreted proteins, many of which are peptidases (including subtilisins and carboxypeptidases) and chitinases[9]. Some of these enzymes appear to have been acquired via horizontal gene transfer (e.g. bacterial GH30 chitinases) and duplicated specifically in P. chlamydosporia, highlighting its evolutionary specialization for nematode parasitism[9]. In summary, P. chlamydosporia carries an extensive enzymatic arsenal (proteases, glycosidases, deacetylases, etc.) that together enable it to breach nematode eggs and exploit them as a nutrient source[2][5].
Induced Systemic Resistance and Plant Growth Promotion#
Beyond direct parasitism, P. chlamydosporia can interact positively with plants. It readily colonizes roots endophytically, and some studies show it can prime plant defenses and promote growth. For instance, colonization of tomato roots by a strain of P. chlamydosporia (PC-170) significantly stimulated plant growth and vigor[5]. In split-root experiments, treated plants grew larger and had fewer nematode galls than untreated controls, even in the uncolonized half of the root system. Molecular analyses revealed that P. chlamydosporia activates jasmonic acid (JA) and salicylic acid (SA) defense pathways in the host. Key marker genes for JA- and SA-mediated defenses were strongly induced in tomato roots after fungal colonization[5]. The result is an induced systemic resistance (ISR) state: systemic plant resistance to nematodes (and even some foliar pathogens) was enhanced by root colonization[5].
Similarly, a different study found that certain P. chlamydosporia isolates could trigger tomato defenses to reduce M. incognita infection, with activation of the SA pathway (PR-1 gene) and the JA pathway (LOX D gene) observed in planta[6]. These defense signals likely contribute to limiting nematode establishment. In some cases, the fungus’ endophytic presence alone (without nematodes present) is enough to upregulate defense genes. Overall, P. chlamydosporia’s dual role as a root colonizer and egg parasite means it can act both as a localized
In practical terms, plants treated with P. chlamydosporia often show better gbiopesticide and as a biostimulant of plant immunity[5][6].rowth and stress tolerance. Field observations and greenhouse studies have noted increased root biomass, nutrient uptake, and resistance to soil-borne diseases in inoculated plants. This is thought to result from a combination of nutrient provision (from parasitized nematode biomass), hormonal interactions, and elicitation of plant defenses. Thus, P. chlamydosporia not only reduces pest pressure but can contribute to overall plant health.
Formulation and Field Application Strategies#
To apply P. chlamydosporia in agriculture, researchers and companies have developed various formulations and delivery methods. The fungus can be formulated as dry granules, liquids, or spores, often mixed with carriers or other beneficial microbes. For example, one commercial product contains P. chlamydosporia at ~10^8 CFU/g together with the nematophagous fungus Arthrobotrys oligospora and a beneficial Glomus or soil drench of spore suspensions, seed or root coating with conidial formulations, and broamycorrhiza. This bioformulation is approved for organic farming and is applied 2–3 times per crop at about 2 L/ha via irrigation or drip systems[3]. It can also be used as a root-dip treatment for seedlings before transplanting.
Field application typically involves delivering the fungus to the root zone where nematodes lay eggs. Common strategies include drip irrigation dcasting granules into soil. In drip-irrigated greenhouses, P. chlamydosporia can be introduced in the irrigation water early in the season, achieving uniform root colonization. For open-field crops without irrigation systems, researchers have experimented with simple tractor-mounted sprayers or granular application to inoculate large areas without high-tech equipment[3]. Some studies have also trialed combining P. chlamydosporia with organic amendments (e.g. compost) or supplements like chitosan to boost its survival and activity. For instance, adding chitosan to soil has been shown to enhance fungal infection of nematode eggs by stimulating the fungus’s enzymatic activity.
Timing and frequency of application are important. Early season applications (before nematode peaks) allow the fungus to establish and encounter nematode eggs over the cropping cycle. Many protocols recommend multiple applications at critical crop stages. Formulations are designed for shelf stability; in practice, producers store lyophilized spores or granules that can be rehydrated. However, ensuring high field efficacy remains challenging, as discussed below.
Challenges and Future Outlook#
Although P. chlamydosporia has clear potential as an eco-friendly nematode biocontrol agent, several challenges limit its widespread adoption.
- Environmental factors can greatly influence its performance: high levels of readily-available carbon (e.g. from organic matter) or unfavorable soil pH can suppress the fungus’s enzyme production and parasitic activity[5]. For example, lab studies found that glucose initially represses but later stimulates VCP1 production, while acidic pH generally reduces VCP1 levels[5]. Such sensitivities mean that soil conditions (nutrient status, pH, moisture) must be conducive for the fungus to thrive.
- Strain selection and consistency are also issues. As noted, different isolates vary in virulence, host range, and enzyme expression[7][1]. Identifying robust, broadly effective strains requires screening and genetic analysis. Modern genomics is aiding this process: the genome and transcriptome of P. chlamydosporia have revealed many candidate virulence genes (e.g. secreted GH30 chitinases, proteases under positive selection) that could serve as markers for efficacy[9]. Gene expression studies under different conditions are illuminating how to optimize culture and formulation conditions.
- Formulation and application technology remain hurdles. Reviews note that “a lack of good formulations with consistent field efficacy” has hindered commercialization of endophytic nematode biocontrol agents[3]. Ensuring shelf stability, ease of use, and uniform field distribution is complex. For example, drying or encapsulation methods must keep fungal propagules viable. UV radiation, temperature swings, or antagonistic soil microbes can also reduce field persistence. Hence, future R&D is focusing on protective formulations (e.g. oil dispersions, nano-carriers) and co-formulants (e.g. humic substances) to improve survival. Beyond formulation, broader challenges include regulatory hurdles (registration of microbial pesticides), integration into cropping systems, and education of growers. Climate change may increase nematode pressures, heightening demand for biocontrol solutions. On the upside, P. chlamydosporia’s ability to act as an endophyte fits well with sustainable agriculture: it can work in IPM programs alongside cover crops, solarization, and reduced chemicals. Continued field trials and farmer-partnered projects are needed to establish best practices. Looking ahead, research is expanding on P. chlamydosporia. Genomic tools have opened new avenues: for instance, Lin et al. (2018) assembled the fungus’s chromosomes and compared strains from different nematodes, revealing unique gene duplications and horizontal gene transfers linked to parasitism[9]. Such insights could guide strain improvement or even genetic engineering to enhance biocontrol traits. Scientists are also exploring synergies with other biocontrols (e.g. combining P. chlamydosporia with nematicidal bacteria or fungi) and with biostimulants like chitosan[8][2]. Ultimately, overcoming the current challenges will depend on a detailed understanding of the plant-fungus-nematode system and on tailoring applications to local conditions[1][3].
Spotlight on Research: Pochonia chlamydosporia – Lin et al. (2018)#
Brief Overview#
Lin et al. (2018) performed a comprehensive genomic and secretomic analysis of P. chlamydosporia[9]. They sequenced the genome of strain PC170 (isolated from root-knot nematode eggs) and compared it with strain PC123 (from cyst nematode eggs) to explore genetic differences related to host specificity. Using bioinformatics, they identified putative chromosomes and annotated all predicted proteins. A special focus was on the secretome – the set of proteins secreted by the fungus – since these are key to infection. The study involved collaborators from Beijing Normal University, the Chinese Academy of Agricultural Sciences (Institute of Vegetables and Flowers), and the University of Alicante in Spain.
Key Insights#
The study uncovered ~1,750 secreted proteins in P. chlamydosporia, many of which are enzyme families implicated in parasitism. The secretome is rich in subtilisin-like proteases, carboxypeptidases, and various glycoside hydrolases (e.g. chitinases)[9]. Comparative analyses highlighted gene duplications and divergence: for example, two duplicated gene pairs encoding GH30 glycosyl hydrolases (enzymes that degrade chitosan) were identified in strain PC170. These GH30 genes appear to have been acquired by horizontal transfer from bacteria and are absent in many insect-pathogenic fungi, suggesting they are specialized adaptations of nematophagous fungi[9].
Additionally, the authors found evidence of positive selection acting on certain secreted proteins, indicating rapid evolution of virulence factors. Transcriptome comparisons under nutrient stress showed that P. chlamydosporia upregulates proteases and carbohydrate-active enzymes when resources are limited, reflecting a strategy to exploit host egg nutrients[9]. In essence, Lin et al. provided a genomic blueprint of P. chlamydosporia’s infection machinery, linking genetic features to its egg-parasitic lifestyle.
Why This Matters#
This research is significant because it elucidates the molecular basis of P. chlamydosporia’s biocontrol ability. By cataloging its secretome and identifying unique enzyme genes (e.g. the bacterial-derived GH30 chitosanases), the study points to specific targets for enhancing efficacy. For instance, the GH30 enzymes may be critical for breaking down nematode eggshells, so strains expressing them strongly could be superior biocontrol agents. The finding of positively-selected genes also hints at which proteins may govern host preference or virulence. Practically, this knowledge can inform strain selection: molecular markers from these genes could help screen for highly virulent isolates. On a broader level, understanding the genome aids efforts to formulate better products (e.g. by knowing what environmental triggers induce key enzymes).
Moreover, the collaborative nature of the work – combining Chinese and Spanish expertise – highlights the global interest in sustainable nematode management. It sets a benchmark for future functional studies, such as knocking out or overexpressing candidate genes to test their roles. The study’s publication in Scientific Reports (a Nature Research journal) and its extensive dataset (including a free PMC article) make it a valuable resource for nematologists and biocontrol developers.
Summary Table: Spotlight Study#
| Category | Details |
| Lead Researchers | Runmao Lin et al.[9] |
| Affiliations | Beijing Normal Univ.; Institute of Vegetables & Flowers (CAAS), Beijing; Univ. of Alicante, Spain |
| Research Focus | Genomic and secretome analysis of P. chlamydosporia |
| Key Breakthroughs | – Assembled 7 putative chromosomes of strain 170. – Identified ~1,750 secreted proteins (rich in proteases and chitinases). – Discovered two GH30 chitosanase gene pairs likely acquired by horizontal transfer. – Detected 32 secreted proteins under positive selection. |
| Collaborative Efforts | SiChina–Spain institutions) |
| Published Work | Scientific Reports (Nat. Publ. Gr.) |
| Perspective | Scientific Reports (Nat. Publ. Gr.) |
| Publication Date | Jano-Spanish collaboration (n 2018 |
| Location | Beijing (China) |
| Key Findings | – Secretome is loaded with enzymes (subtilisins, chitinases) for nematode egg parasitism[9]. – Unique GH30 chitosanases (horizontal gene transfer) suggest special adaptation to eggshell degradation[9]. – Insights into gene evolution and potential virulence factors for nematode control. |
Conclusion#
Pochonia chlamydosporia exemplifies an eco-friendly approach to nematode control by combining direct parasitism with plant-beneficial traits. Its global distribution, multitrophic lifestyle, and arsenal of lytic enzymes make it well-suited to target root-knot and cyst nematodes while also stimulating plant defenses[2][5]. Numerous studies demonstrate its ability to reduce nematode populations and improve crop health in greenhouse and field trials[4][3]. Formulations based on P. chlamydosporia are compatible with organic farming and integrated pest management.
Continued research – from genomics to agronomic trials – is refining how best to deploy this fungus. Challenges remain (such as formulation stability and environmental variability), but advances in understanding its genetics and interaction with plants are opening new possibilities[9][3]. As chemical nematicides are phased out, P. chlamydosporia stands out as a biopesticide of the future, harnessing nature’s own checks on pests to protect crops sustainably.
References#
- Manzanilla-López RH, Esteves I, Finetti-Sialer MM, Hirsch PR, Ward E, Devonshire J, Hidalgo-Díaz L (2013) Pochonia chlamydosporia: Advances and challenges to improve its performance as a biological control agent of sedentary endo-parasitic nematodes. J. Nematol. 45(1):1–7journals.flvc.orgjournals.flvc.org.
- Aranda-Martínez A, Lenfant N, Escudero N, Zavala-González EA, Henrissat B, López-Llorca LV (2016) CAZyme content of Pochonia chlamydosporia reflects that chitin and chitosan modification are involved in nematode parasitism. Environ. Microbiol. 18(11):4200–4215pubmed.ncbi.nlm.nih.govpubmed.ncbi.nlm.nih.gov.
- Sellitto VM, Curto G, Dallavalle E, Ciancio A, Colagiero M, Pietrantonio L, Birăescu G, Stoleru V, Storari M (2016) Effect of Pochonia chlamydosporia-based formulates on regulation of root-knot nematodes and plant growth response. Front. Life Sci. 9(3):[Full Text]researchgate.net.
- Yang JI, Loffredo A, Borneman J, Becker JÓ (2012) Biocontrol efficacy among strains of Pochonia chlamydosporia obtained from a root-knot nematode suppressive soil. J. Nematol. 44(1):67–71pubmed.ncbi.nlm.nih.gov.
- Ward E, Kerry BR, Manzanilla-López RH, et al. (2012) The Pochonia chlamydosporia serine protease gene vcp1 is subject to regulation by carbon, nitrogen and pH: Implications for nematode biocontrol. PLoS ONE 7(4):e35657pubmed.ncbi.nlm.nih.govpubmed.ncbi.nlm.nih.gov.
- Ghahremani Z, Escudero N, Sorribas FJ, et al. (2019) Pochonia chlamydosporia induces plant-dependent systemic resistance to Meloidogyne incognita. Front. Plant Sci. 10:945frontiersin.orgfrontiersin.org.
- Morton, C. O., Hirsch, P. R., Peberdy, J. P., & Kerry, B. R. (2003). Cloning of and genetic variation in protease VCP1 from the nematophagous fungus Pochonia chlamydosporia. Mycological Research, 107(1), 38–46. https://doi.org/10.1017/S0953756202007050
- Suárez-Fernández M, Sambles C, Lopez-Moya F, et al. (2021) Chitosan modulates Pochonia chlamydosporia gene expression during nematode egg parasitism. Environ. Microbiol. 23(9):4980–4997pubmed.ncbi.nlm.nih.govpubmed.ncbi.nlm.nih.gov.
- Lin R, Qin F, Shen B, et al. (2018) Genome and secretome analysis of Pochonia chlamydosporia provide new insight into egg-parasitic mechanisms. Sci. Rep. 8:1123pubmed.ncbi.nlm.nih.govpubmed.ncbi.nlm.nih.gov.