Photorhabdus luminescens: Nature’s Bioluminescent Insect Pathogen and Biopesticide Agent
Overview of the Microbe Photorhabdus luminescens is a Gram-negative, rod-shaped bacterium in the family Morganellaceae (formerly Enterobacteriaceae) that is notable for its bioluminescence and insect-pathogenic...
- 1
Uses and Actions of Photorhabdus Bacteria
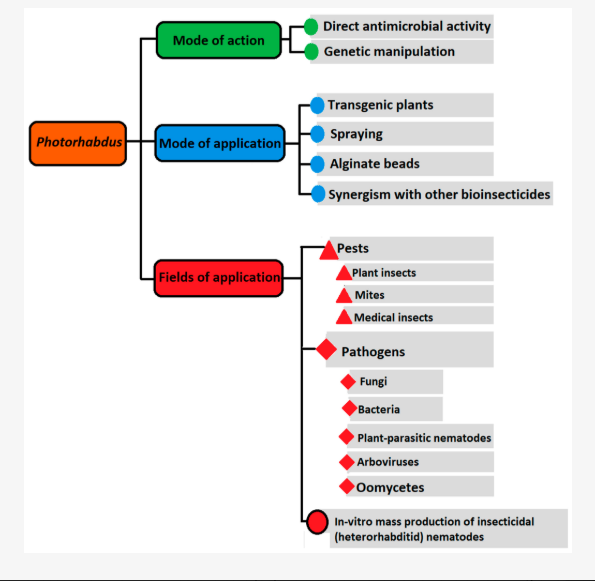
This diagram shows how Photorhabdus bacteria work, how they can be applied, and where they are useful
Overview Table of Photorhabdus luminescens
- Feature
Description
- Scientific Name
Photorhabdus luminescens
- Classification
Phylum: Pseudomonadota; Class: Gammaproteobacteria; Order: Enterobacterales; Family: Morganellaceae
- Habitat
Symbiotic in the gut of Heterorhabditid nematodes; released into insect haemocoel upon infection
- Key Functions
Rapid insect killing; cadaver bioconversion; antibiotic production
- Notable Abilities
Production of high-molecular-weight toxin complexes (Tca), Mcf toxin; secretion of degradative enzymes
- Applications
Biopesticide in agriculture; source of novel bioactive compounds; model for symbiosis research
- Genetic Engineering Potential
Targets: mcf, tca loci; Tools: CRISPR/Cas9 for attenuated strains
- Challenges
Scale-up of fermentation; environmental safety and non-target impacts; regulatory approval
- Future Prospects
Engineering toxin-free delivery vehicles; bioluminescent biosensors; integration in circular-bioeconomy
Overview of the Microbe#
Photorhabdus luminescens is a Gram-negative, rod-shaped bacterium in the family Morganellaceae (formerly Enterobacteriaceae) that is notable for its bioluminescence and insect-pathogenic lifestyle[1]. It is one of several Photorhabdus species (including P. temperata and P. asymbiotica) that live in a mutualistic symbiosis with nematodes of the genus Heterorhabditis[1]. In nature, P. luminescens is carried by infective juvenile (IJ) nematodes into the hemocoel of insect larvae. Upon release into the insect’s blood, the bacteria proliferate rapidly and produce a suite of toxins and enzymes that kill the host within 24 hours[2][1]. The resulting insect cadaver becomes a nutrient-rich soup of bacterial and nematode biomass; P. luminescens secretes compounds that prevent invasion by other microbes during this phase[2][3].In laboratory culture P. luminescens displays a characteristic phase variation: a primary form that glows blue-green (due to the luxCDABE luciferase operon) and secretes secondary metabolites, versus a secondary form that is non-luminescent and has lost many toxin and pigment genes[1][5]. The genome of P. luminescens is unusually rich in genes for secondary metabolites: dozens of gene clusters encode toxins, antibiotics, enzymes, and insecticidal proteins[3][5]. This metabolic arsenal makes the bacterium highly pathogenic and adaptable. P. luminescens and its nematode host have a worldwide distribution, matching the habitats of Heterorhabditis spp. (found from temperate to tropical soils)[3]. Because it is specific to insects and survives poorly above ~34 °C, P. luminescens poses little risk to humans in the wild (unlike its relative P. asymbiotica, which can infect humans)[1].

Entomopathogenic Symbiosis#
The defining biological partnership of P. luminescens is its mutualism with Heterorhabditis nematodes. In this symbiosis, nematodes serve as vectors that deliver the bacteria into insect hosts, and the bacteria provide nutrients and antimicrobial defense for the nematode’s offspring. Specifically, the infective juvenile (IJ) of Heterorhabditis carries P. luminescens cells in its gut lumen in a nonmotile monoculture[2]. When the IJ enters an insect larva (typically through natural openings or by penetrating the cuticle), it regurgitates the bacteria into the insect hemocoel in response to insect hemolymph signals[2][1]. The bacteria multiply exponentially and secrete potent toxins that kill the insect within about one day[2][1]. This rapid insect mortality (a 50% lethal dose of fewer than 30 bacterial cells has been reported) is due to both toxins and degradative enzymes[2]. As the insect tissues are liquefied, nematode juveniles feed on the growing mass of bacteria and cadaver fluids. Meanwhile P. luminescens produces antibiotics, anti-fungal compounds, and other bactericidal products that protect the cadaver from colonization by competing microbes[2][3]. After several nematode generations, new infective juveniles emerge from the depleted cadaver, now colonized internally by P. luminescens, and the cycle can repeat[2][1].
This tripartite lifestyle is remarkably efficient: the nematode propagates only when partnered with a P. luminescens strain it recognizes, and the bacterium only gets released when needed to exploit an insect host. The collaboration is species-specific and highly coevolved[2]. Notably, P. luminescens is so virulent that it can kill a wide range of insect hosts (including agriculturally important pests like caterpillars and beetle larvae) even when introduced by itself. Because of this, the nematode–bacterium complex is studied and applied as a biological control agent. For example, P. luminescens strain TT01 has been commercialized in combination with Heterorhabditis bacteriophora nematodes (e.g. product Xenorhabdus-based insecticide). In nature, this system often colonizes below-ground pests (e.g. grubs), but it also has potential against foliar pests if formulations can deliver bacteria or nematodes to foliage or soil[3].
Toxin Complexes and Insecticidal Mechanisms#
A hallmark of P. luminescens biology is its rich repertoire of insecticidal toxins. Genomic and biochemical studies have grouped these toxins into several families[1][3]. The major toxin classes are:
- Toxin complexes (Tc proteins): These are giant multi-subunit assemblies (composed of TcA, TcB, TcC, Tcd, etc.) that function like molecular syringes. A Tc complex can bind to insect gut or blood cells and inject a toxic enzyme inside. P. luminescens Tc toxins cause broad-spectrum mortality, even when ingested, by targeting cell cytoskeleton (e.g. glycosylating actin) and other processes[1][8]. Four major Tc operons (Tca, Tcb, Tcc, Tcd types) have been identified in P. luminescens genomes[1]. Each operon encodes multiple protein chains that assemble into a large (>2 MDa) complex.
- Photorhabdus insect-related (Pir) toxins: These are smaller binary (two-component) toxins, related to Bacillus thuringiensis Cry toxins. Each PirA/PirB pair must be ingested to become active and can perforate insect gut cells. Insects exposed to Pir toxins show gut paralysis and death. Pir toxins have a δ-endotoxin-like structure and are lethal to certain mosquito and moth larvae.
- Makes Caterpillars Floppy (Mcf) toxins: Mcf toxins (notably Mcf1) are proteins that induce widespread apoptosis in insect tissues. They were named for causing larvae to go “floppy” (loss of body turgor) before dying. Mcf1 contains multiple domains – including a BH3-like motif that mimics pro-apoptotic factors – and requires host uptake. Recent cryo-EM studies have revealed that Mcf1 is a “seahorse-shaped” molecule with a long tail and a multi-domain head[8]. When P. luminescens injects Mcf1 into an insect cell, the tail initially binds to the cell membrane and undergoes a conformational change upon interacting with the host’s ARF3 protein. This triggers three autocleavage events in Mcf1, releasing two internal toxic domains into the cell cytoplasm[8]. These effectors then disrupt essential host processes, causing rapid cell death and immune evasion. The structural details of Mcf1 elucidated by Belyy et al. help explain how this single-chain toxin folds and activates within an insect host[8].
- Photorhabdus Virulence Cassettes (PVC) and other effectors: PVCs are phage tail–like assemblies that inject effector proteins into insect cells. Other smaller toxins include Photox (an ADP-ribosyltransferase), PaxAB binary toxins, Pit toxins, and various enzymes (proteases, lipases) that degrade insect tissues. In total, P. luminescens carries pathogenicity islands with dozens of toxin genes[3][1].
In sum, P. luminescens employs a multipronged assault on insects: membrane-perforating complexes (Tcs), gut toxins (Pir, Cry-like), apoptotic toxins (Mcf), and degradative enzymes. The combined effect is a very potent, broad-spectrum biocontrol activity. Importantly, many of these toxin families have no homologs in mammals, giving them high specificity for invertebrates, which underlies interest in their use as organic pesticides[8].
Antibiotic Production and Microbial Competition#
Beyond killing insects, P. luminescens must secure the nutrient-rich cadaver from other microbes. It does so by producing diverse antibiotics and bacteriocins during its reproductive phase[3][5]. These secondary metabolites inhibit bacteria and fungi that might otherwise consume the insect corpse. For example, P. luminescens produces a stilbene antibiotic (3,5-dihydroxy-4-isopropyl-trans-stilbene, or ST) that has dual functions: it suppresses the insect’s melanization response by inhibiting the host enzyme phenoloxidase, and it acts as a broad-spectrum antibiotic against invaders[4]. Eleftherianos et al. showed that mutant bacteria unable to make ST are less virulent in caterpillars and allow more contaminating microbes to grow in the cadaver[4]. In addition to ST, P. luminescens synthesizes various polyketides, nonribosomal peptides, and bacteriocins. Screening of a South African strain (Photorhabdus sp. ETL) found it produces xenorhabdins, xenocoumacins, nematophin (an indole alkaloid), genistein (isoflavone), anthraquinones, chitinases, and more[5]. All of these have known or suspected antimicrobial activity. In effect, P. luminescens sterilizes the insect milieu to ensure its nematode partner can reproduce in peace[3][5]. This antibiotic arsenal has attracted interest for discovery of new drugs as well as for engineering of enhanced biopesticide strains.
Bioluminescence and Biosensing Applications#
A unique trait of P. luminescens is its bioluminescence: primary-phase cells harbor the luxCDABE operon, which encodes a luciferase that catalyzes light emission at ~490 nm[1]. In fact, infected insect larvae can literally glow in the dark as the bacteria multiply. The precise ecological function of luminescence in P. luminescens remains unclear, but biologically it is a useful marker. For biotechnology, the luciferase from P. luminescens is especially valuable because it is more thermostable than that of marine luminescent bacteria (e.g. Vibrio fischeri) and does not require the special salts that Vibrio needs[7].
As a result, the P. luminescens lux genes have become standard tools in biosensor and reporter design. In the simplest case, the entire luxCDABE operon can be transferred to other bacteria, yielding strains that emit light constitutively or in response to a promoter of choice. For example, Prest et al. (1997) fused the P. luminescens lux operon to an E. coli nitrate-inducible promoter (narG), creating a nitrate biosensor that could detect levels down to 5×10^-5 M[6]. In environmental monitoring, lux-based whole-cell assays are popular because they provide a real-time optical readout: samples that are toxic will reduce bacterial luminescence. Sihtmäe et al. (2011) demonstrated that E. coli strains engineered with the P. luminescens lux genes are effective biosensors for both organic chemicals and heavy metals[7]. In their Flash-Assay format, these lux E. coli were as sensitive as the standard Vibrio fischeri Microtox test for pollutants like Hg, Cd, aniline, etc[7]. Because P. luminescens luciferase functions well at 30–40 °C, these biosensors work at common laboratory temperatures where V. fischeri would be suboptimal[7].
In short, P. luminescens’s bioluminescence is not just a curiosity but a practical resource. Its lux operon is widely used to tag pathogens in infection studies, to create bio-reporter strains for drug screening, and to develop in vivo imaging systems. Researchers have also used P. luminescens itself (or its lux mutants) as indicator bacteria in soil and water assays. The ease of measuring bacterial light output makes these tools “self-reporting” – no external substrates are needed, and responses are continuous. Thus, P. luminescens has lit the path for a generation of living biosensors that help detect environmental toxins and monitor microbial processes[6][7].
Challenges and Future Potential#
Despite its promise as a biopesticide, P. luminescens faces several challenges for practical deployment. One issue is cost-effective production and formulation. Mass-rearing of bacteria (or of their nematode hosts) must be economically viable. P. luminescens has complex nutritional needs, and maintaining the virulent “primary” phase in large-scale culture can be difficult, since spontaneous phase variation can yield non-luminescent, less-virulent secondary cells[5]. At least one agricultural review notes that field use has been slow due to production costs, uneven distribution of nematode hosts in the environment, and trait stability during storage and culture[3]. However, progress is being made: research has demonstrated cheaper media formulations for Photorhabdus/ Heterorhabditis cultures, and advances in fermentation technology are enabling larger batches with consistent quality[3.
Another concern is safety and regulation. Although P. luminescens cannot grow at human body temperature, its close relative P. asymbiotica has caused occasional human infections. Therefore, any use of P. luminescens as a biocontrol agent must be carefully evaluated for off-target effects and ecological impact. Regulatory agencies require demonstration that released bacteria (or their toxins) will not harm non-target insects, beneficial soil fauna, or vertebrates. Long-term environmental studies are still needed to address these questions.
On the other hand, P. luminescens also offers many future opportunities. Its genome has been sequenced and mined for novel bioactive compounds, and new Photorhabdus species and strains continue to be discovered in various parts of the world[3]. Synthetic biology approaches could harness specific toxin genes (e.g. expressing Pir or Mcf in transgenic crops) or tweak bacterial regulators to enhance safety and potency. The structural insights into toxins (like the recent Mcf1 structure[8]) pave the way for protein engineering: one could imagine designing chimeric toxins with new target specificities or environmental stabilities.
Moreover, P. luminescens might contribute beyond pest control. Its antibiotics have potential as leads for new antimicrobials in medicine. Its immunomodulatory compounds could inspire novel pharmaceuticals. Researchers are even exploring the delivery of antigens via Photorhabdus-derived systems for vaccines (analogous to how Bt toxins have been repurposed). In sustainable agriculture, P. luminescens could integrate into integrated pest management: for example, its bacteria or purified toxins might be applied alongside beneficial insects or used to prime plant defenses.In summary, while practical hurdles remain (production, regulation, stability), the unique biology of P. luminescens suggests it will have a growing role in biocontrol. Ongoing research and biotech advances continue to unlock its potential, turning nature’s insect pathogen into a tool for environmentally friendly pest management[3][8].
Spotlight on Research: Photorhabdus luminescens#
Brief Overview#
- A recent Nature Communications study by Belyy et al. (2023) has provided the first high-resolution structure of the P. luminescens Mcf1 toxin[8]. Using cryo-electron microscopy, the authors (led by Stefan Raunser’s team) resolved the 3.6 Å structure of Mcf1, revealing its overall “seahorse” architecture with a long tail and a head containing two toxic domains[8]. The analysis showed how the tail undergoes a conformational shift upon binding to a host ADP-ribosylation factor (ARF3), triggering sequential autoproteolysis events that release the toxin’s effector modules inside the insect cell[8]. This work elucidates the molecular mechanism of Mcf1 activation at an unprecedented level.
Key Insights#
- First-ever Mcf1 structure: Mcf1’s tail and head domains were visualized, showing how each is arranged and how the toxin changes shape upon activation[8].
- Seahorse-like mechanism: The “tail” contains putative membrane-binding and translocation domains, while the “head” houses two effectors, an activator-binding domain, and an internal autoprotease[8].
- Host-triggered activation: Binding of host ARF3 to the tail relocates the toxin’s own cleavage site into its protease, causing three self-cleavage cuts and liberation of the toxic payloads[8].
- Conservation with human pathogens: The structural theme (tail-hook activation) is similar to toxins from Clostridioides difficile, suggesting a broader relevance across bacteria[8].
Why This Matters#
Understanding how Mcf1 works at the molecular level has important implications. By revealing the steps that lead to insect cell death, this study provides blueprints for engineering improved biocontrol agents. For instance, synthetic biologists could modify Mcf1’s recognition or payload domains to target specific pest species or to evade known resistance. Additionally, the concept of a “hidden” autocleavage activation could inspire new designs of toxin-based delivery systems for agricultural or medical use. More broadly, this work exemplifies how fundamental structural biology can inform applied science: deciphering a bacterial weapon illuminates strategies to harness or neutralize it.
Summary Table: Spotlight Study#
| Category | Details |
| Lead Researchers | Alexander Belyy (1st author), Philipp Heilen, Philine Hagel, Oliver Hofnagel, Stefan Raunser (senior) |
| Affiliations | Max Planck Institute for Molecular Physiology, Dortmund, Germany |
| Research Focus | Structural biology of Mcf1 toxin from P. luminescens |
| Key Breakthroughs | First cryo-EM structure of Mcf1; detailed activation mechanism (ARF3-triggered autoproteolysis releasing effectors) |
| Collaborative Efforts | MPI Dortmund (Raunser lab); collaborators in structural biology |
| Published Work | Belyy et al., Nat. Commun. 14:8226 (2023)[8] |
| Perspective | Provides molecular insight into an insecticidal toxin; informs design of new biopesticides |
| Publication Date | 12 Dec 2023 |
| Location | Dortmund, Germany |
| Key Findings | Mcf1 is a seahorse-shaped protein: tail binds host membrane and ARF3, triggering cleavage and release of two toxin domains[8] |
Conclusion#
Photorhabdus luminescens exemplifies nature’s dual gift of microbial novelty and pest control potential. Its biology bridges fundamental microbiology and practical agriculture: as a bacterium it offers insights into symbiosis, toxin evolution, and antibiotic discovery; as a biocontrol agent it contributes to safer, more sustainable pest management. The organism’s remarkable arsenal of toxins and antibiotics arises from an evolutionary niche—living inside nematodes and insects—that has honed its genome into an “all-in-one” pest weapon[2][1]. Although challenges remain in harnessing P. luminescens at scale, recent research (e.g., the Mcf1 structure) shows that scientific advances continue to unlock its secrets. In the coming years, P. luminescens and its products are likely to play an increasing role in integrated pest management, serving both as direct biopesticides (through formulated nematodes or bacterial preparations) and as sources of novel bio-insecticides and biosensor tools. Through careful development and stewardship, this glowing insect pathogen can help reduce reliance on chemical pesticides, protect crops, and inspire new biotechnologies for a greener future.
References#
- Rodou A, Ankrah DO, Stathopoulos C. 2010. Toxins and secretion systems of Photorhabdus luminescens. Toxins 2(6):1250–1264.
- Ciche TA, Ensign JC. 2003. For the insect pathogen Photorhabdus luminescens, which end of a nematode is out? Appl. Environ. Microbiol. 69(4):1890–1897.
- Abd-Elgawad MM. 2021. Photorhabdus spp.: An overview of the beneficial aspects of mutualistic bacteria of insecticidal nematodes. Plants 10(8):1660.
- Eleftherianos I, Boundy S, Joyce SA, Aslam S, Marshall JW, Cox RJ, Simpson TJ, Clarke DJ, ffrench-Constant RH, Reynolds SE. 2007. An antibiotic produced by an insect-pathogenic bacterium suppresses host defenses through phenoloxidase inhibition. Proc. Natl. Acad. Sci. USA 104(7):2419–2424.
- Lulamba TE, Green E, Serepa-Dlamini MH. 2021. Photorhabdus sp. ETL antimicrobial properties and characterization of its secondary metabolites by gas chromatography–mass spectrometry. Life (Basel) 11(8):787.
- Prest AG, Winson MK, Hammond JR, Stewart GS. 1997. The construction and application of a lux-based nitrate biosensor. Lett. Appl. Microbiol. 24(5):355–360.
- Kurvet I, Ivask A, Bondarenko O, Sihtmäe M, Kahru A. 2011. LuxCDABE–Transformed constitutively bioluminescent Escherichia coli for toxicity screening: comparison with naturally luminous Vibrio fischeri. Sensors 11(8):7865–7878.
- Belyy A, Heilen P, Hagel P, Hofnagel O, Raunser S. 2023. Structure and activation mechanism of the Makes caterpillars floppy 1 toxin. Nat. Commun. 14:8226.