Pseudomonas fluorescens: A Multitalented Plant-Associated Bacterium
Bacillus subtilis is a robust, Gram-positive bacterium widely recognized for its adaptability and efficiency in various environments.
- 1
How Pseudomonas fluorescens 2P24 Removes Nitrogen from Wastewater
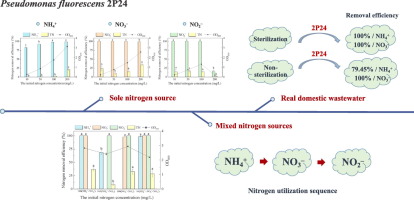
This figure shows how the bacterium Pseudomonas fluorescens 2P24 efficiently removes different nitrogen compounds—like ammonia (NH₄⁺), nitrite (NO₂⁻), and nitrate (NO₃⁻)—from both clean and real domestic wastewater.
Overview Table of Pseudomonas fluorescens
- Feature
Description
- Scientific Name
Pseudomonas fluorescens (Flügge 1886) Migula 1895
- Classification
Phylum: Pseudomonadota; Class: Gammaproteobacteria; Order: Pseudomonadales; Family: Pseudomonadaceae
- Habitat
Soil, plant rhizosphere, freshwater, phyllosphere
- Key Functions
Plant growth promotion; biocontrol; pollutant degradation
- Notable Abilities
Siderophore production; biofilm formation; broad substrate utilisation
- Applications
Biofertilisers; biopesticides; bioremediation; enzyme production
- Genetic Engineering Potential
Plasmid-borne metabolic pathways; CRISPR/Cas editing; fluorescent reporter systems
- Challenges
Environmental variability; inconsistent field efficacy; regulatory frameworks
- Future Prospects
Synthetic biology chassis; AI-guided strain optimisation; integrated biofertiliser–bioremediation systems
Overview of the Microbe#
Pseudomonas fluorescens is a non‑spore‑forming, motile, Gram‑negative rod commonly isolated from soil, water, and plant roots[1]. It secretes the fluorescent siderophore pyoverdine, which gives its name (“fluorescens”) and facilitates iron uptake in iron‑limiting environments. Not typically a human pathogen, it instead thrives as an environmental bacterium with a genome size ranging from ~6.0 to 7.1 Mbp, encoding over 5 500 protein‑coding genes that confer metabolic versatility[2][3].
Genomic sequencing of key strains—such as Pf‑5, SBW25, and Pf0‑1—has revealed a rich complement of catabolic enzymes, efflux pumps, regulatory circuits, and biosynthetic gene clusters for antibiotics and biosurfactants[7][14]. Comparative phylogenomics shows that isolates labeled “P. fluorescens” form a polyphyletic complex, leading to its reclassification as a species complex rather than a single clade[6].

Nutrient-Rich for Food and Feed#
Plant Growth Promotion#
Certain P. fluorescens strains function as plant‑growth‑promoting rhizobacteria (PGPR), enhancing seed germination, root elongation, and nutrient acquisition through multiple mechanisms[9]. They secrete siderophores (e.g., pyoverdine) that solubilize iron, making it available to plants under iron‑limiting soil conditions PMC. Other PGPR traits include phosphate solubilization, indole‑3‑acetic acid (IAA) production, and nitrogen fixation, which collectively improve plant biomass and yield—for example, groundnut yield increased by up to 30 % upon P. fluorescens inoculation[11].
Induced Systemic Resistance and Biocontrol#
P. fluorescens triggers induced systemic resistance (ISR) in plants, priming them to defend against foliar and soilborne pathogens[8]. Key biocontrol metabolites include 2,4‑diacetylphloroglucinol (2,4‑DAPG), pyoluteorin, and cyclic lipopeptides (e.g., viscosin), which inhibit fungi, oomycetes, and bacteria[5][4]. These compounds not only suppress pathogens directly but also act as signals modulating gene expression within the rhizosphere microbiome, enhancing community‑level disease suppression[4].
Biodegradation and Bioremediation#
Organic Pollutant Degradation#
Many P. fluorescens strains degrade environmental contaminants—including polycyclic aromatic hydrocarbons, chlorinated compounds, and pesticides—via mono‑ and dioxygenases, hydrolases, and peroxidases[12]. For instance, strain ATCC 13525 can utilize polyester polyurethane as a sole carbon and energy source, offering routes to plastic waste valorization[13]. Engineered strains expressing catabolic pathways (e.g., 2,4‑dinitrotoluene degradation) further expand its biodegradation repertoire[15].
Heavy‑Metal and Xenobiotic Remediation#
Some isolates bioaccumulate or transform heavy metals—such as cadmium and arsenic—through biosorption by cell walls, extracellular polymeric substances, and metallophore production, achieving removal efficiencies up to 60 % in contaminated waters[14]. These capabilities make P. fluorescens a candidate for bioremediation of mixed polluted sites.
Secondary Metabolites and Antimicrobial Activity#
P. fluorescens synthesizes a spectrum of secondary metabolites with antimicrobial properties. The phenazine phloroglucinols (e.g., 2,4‑DAPG) suppress soilborne pathogens, while pyoluteorin and pyrrolnitrin target fungal pathogens. Cyclic lipopeptides—such as viscosin and amphisin—serve as biosurfactants and antimicrobials, disrupting membrane integrity of competing microbes[5]. Interspecies signaling studies demonstrate that subinhibitory concentrations of pyoluteorin modulate 2,4‑DAPG biosynthesis via TetR‑family regulators (PhlF, PhlH), highlighting ecological roles beyond direct antibiosis[4].
Phylogenetics and Genetic Engineering Potential#
Phylogenomic analyses place P. fluorescens within a complex of over 50 related species, revealing subclade structure and high genomic diversity across environmental and clinical isolates[17][16]. This diversity underlies varied ecological functions and biotechnological traits.Genetic tools—such as broad‑host‑range plasmids, CRISPR/Cas systems, and integrative vectors—enable pathway engineering in P. fluorescens for enhanced biocontrol, biosurfactant production, and pollutant degradation. Attenuated or chassis strains can house heterologous catabolic genes, mitigating biosafety concerns while retaining robust environmental fitness.
Challenges and Future Potential#
Despite its non‑pathogenic status, regulatory and biosafety assessments are necessary before environmental release, particularly given strain‑level variability[10]. Species‑complex taxonomy complicates strain selection, necessitating genome‑based classification for consistent performance[18].
Future directions include synthetic community design—leveraging interspecies signaling mechanisms—and bioreactor‑based containment for safe, high‑throughput applications. Advances in systems biology and metabolic modeling will accelerate the rational design of tailored P. fluorescens strains for agriculture, waste valorization, and green chemistry.
Spotlight on Research: Pseudomonas fluorescens 2P24#
Brief Overview#
Zhang et al. (2025) characterized P. fluorescens 2P24—isolated from wheat rhizosphere in take‑all decline soil—for its interspecies signaling and biocontrol activities[4].
Key Insights#
They discovered that pyoluteorin acts as a signal sensed by TetR repressors PhlF and PhlH, which modulate transcription of the 2,4‑DAPG biosynthetic operon—enhancing ecological adaptability and coordinated antibiotic production[4].
Why This Matters#
Understanding molecular signaling among rhizobacteria unveils strategies to optimize biocontrol consortia, potentially reducing chemical pesticide use and improving sustainable agriculture.
Summary Table: Spotlight Study#
| Category | Details |
| Lead Researchers | Nannan Zhang et al. |
| Affiliations | School of Life Sciences & Institute of Health Sciences and Technology, Anhui University; Lanzhou University; China Agricultural University |
| Research Focus | Interspecies signaling and 2,4‑DAPG regulation |
| Key Breakthroughs | Identification of pyoluteorin as an interspecies signal; characterization of PhlF/PhlH‑mediated regulation of antibiotic biosynthesis |
| Collaborative Efforts | Multi‑institutional Chinese research group |
| Published Work | Microbiology Spectrum |
| Publication Date | 2025 |
| Location | China |
| Key Findings | Pyoluteorin modulates 2,4‑DAPG synthesis via TetR repressors, revealing conserved signaling mechanisms across Pseudomonas spp.[4] |
Conclusion#
Pseudomonas fluorescens exemplifies a multitalented, plant‑associated bacterium whose metabolic breadth spans nutrient cycling, plant promotion, biocontrol, pollutant detoxification, and production of value‑added biochemicals. Its genomic plasticity and amenability to engineering offer vast potential for sustainable agriculture and environmental biotechnology. Continued genome‑guided strain development and ecological studies will unlock new applications while ensuring biosafety and regulatory compliance.
References#
- Scales, B. S., Dickson, R. P., LiPuma, J. J., & Huffnagle, G. B. (2014). Microbiology, genomics, and clinical significance of the Pseudomonas fluorescens species complex, an unappreciated colonizer of humans. Clinical Microbiology Reviews, 27(4), 927–948. https://doi.org/10.1128/CMR.00044‑14
- Paulsen, I. T., Press, C. M., Ravel, J., Kobayashi, D. Y., Myers, G. S. A., Mavrodi, D. V., DeBoy, R. T., Seshadri, R., Ren, Q., Madupu, R., Dodson, R. J., Durkin, A. S., Brinkac, L. M., Daugherty, S. C., Sullivan, S. A., Rosovitz, M. J., Gwinn, M. L., Zhou, L., Schneider, D. J., Cartinhour, S. W., Nelson, W. C., Weidman, J., Watkins, K., Tran, K., Khouri, H., Pierson, E. A., Pierson, L. S. III, Thomashow, L. S., & Loper, J. E. (2005). Complete genome sequence of the plant commensal Pseudomonas fluorescens Pf‑5: Insights into the biological control of plant disease. Nature Biotechnology, 23(7), 873–878. https://doi.org/10.1038/nbt1110
- Taylor, T. B., Silby, M. W., & Jackson, R. W. (2025). Pseudomonas fluorescens. Trends in Microbiology, 33(2), 250–251. https://doi.org/10.1016/j.tim.2024.11.00
- Zhang, N., Zhu, X., Tao, X., Li, J., Tang, Q., Liu, X., Luo, L.-M., He, Y.-X., & Ge, H. (2025). Interspecies signaling modulates the biosynthesis of antimicrobial secondary metabolites related to biological control activities of Pseudomonas fluorescens 2P24. Microbiology Spectrum, 13(3), Article e01886‑24. https://doi.org/10.1128/spectrum.01886‑24
- Suresh, P., Vellasamy, S., & Rajakrishnan, R. (2022). Pseudomonas fluorescens VSMKU3054-mediated induced systemic resistance in tomato against Ralstonia solanacearum. Physiological and Molecular Plant Pathology, 119, Article 101836. https://doi.org/10.1016/j.pmpp.2022.101836
- Gomila, M., Peña, A., Mulet, M., Lalucat, J., & García‑Valdés, E. (2015). Phylogenomics and systematics in Pseudomonas. Frontiers in Microbiology, 6, Article 214. https://doi.org/10.3389/fmicb.2015.00214
- Fortmann‑Grote C, et al. Genome update for Pseudomonas fluorescens isolate SBW25. Microbiol Resour Announc. 2023;12(2):e00637‑22.
- De Vleesschauwer, D., Djavaheri, M., Bakker, P. A. H. M., & Höfte, M. (2008). Pseudomonas fluorescens WCS374r‑induced systemic resistance in rice against Magnaporthe oryzae is based on pseudobactin‑mediated priming for a salicylic acid‑repressible multifaceted defense response. Plant Physiology, 148(4), 1996–2012. https://doi.org/10.1104/pp.108.127878
- David, B. V. (2018). Pseudomonas fluorescens: A plant‑growth‑promoting rhizobacterium (PGPR) with potential role in biocontrol of pests of crops. In Crop Improvement through Microbial Biotechnology (Chapter 10). Elsevier. https://doi.org/10.1016/B978-0-444-63987-5.00010-4
- Wikipedia contributors. Pseudomonas fluorescens. Wikipedia, The Free Encyclopedia. Retrieved July 2025.
- Sah, S., Krishnani, S., & Singh, R. (2021). Pseudomonas mediated nutritional and growth promotional activities for sustainable food security. Current Research in Microbial Sciences, 2, Article 100084. https://doi.org/10.1016/j.crmicr.2021.100084
- Gutiérrez, E. J., Abraham, M. d. R., Baltazar, J. C., Vázquez, G., Delgadillo, E., & Tirado, D. (2020). Pseudomonas fluorescens: A bioaugmentation strategy for oil‑contaminated and nutrient‑poor soil. International Journal of Environmental Research and Public Health, 17(19), Article 6959. https://doi.org/10.3390/ijerph17196959
- Howard, G. T., & Blake, R. C. (1998). Growth of Pseudomonas fluorescens on a polyester–polyurethane and the purification and characterization of a polyurethanase–protease enzyme. International Biodeterioration & Biodegradation, 42(3), 213–220. https://doi.org/10.1016/S0964-8305(98)00051-1
- Riera, N., Davyt, D., Durán, R., Iraola, G., Lemanceau, P., & Bajsa, N. (2023). An antibiotic produced by Pseudomonas fluorescens CFBP2392 with antifungal activity against Rhizoctonia solani. Frontiers in Microbiology, 14, Article 1286926. https://doi.org/10.3389/fmicb.2023.1286926
- Monti, M. R., Smania, A. M., Fabro, G., Álvarez, M. E., & Argaraña, C. E. (2005). Engineering Pseudomonas fluorescens for biodegradation of 2,4-dinitrotoluene. Applied and Environmental Microbiology, 71(12), 8864–8872. https://doi.org/10.1128/AEM.71.12.8864-8872.2005
- Garrido‑Sanz, D., Meier‑Kolthoff, J. P., Göker, M., Martín, M., Rivilla, R., & Redondo‑Nieto, M. (2016). Genomic and genetic diversity within the Pseudomonas fluorescens complex. PLOS ONE, 11(2), Article e0150183. https://doi.org/10.1371/journal.pone.0150183
- Scales, B. S., Erb‑Downward, J. R., Huffnagle, I. M., LiPuma, J. J., & Huffnagle, G. B. (2015). Comparative genomics of Pseudomonas fluorescens subclade III strains from human lungs. BMC Genomics, 16, Article 1032. https://doi.org/10.1186/s12864-015-2261-2
- Gomila, M., Peña, A., Mulet, M., Lalucat, J., & García‑Valdés, E. (2015). Phylogenomics and systematics in Pseudomonas. Frontiers in Microbiology, 6, Article 214. https://doi.org/10.3389/fmicb.2015.00214
- Silva‑Rocha R, et al. Prospects of Pseudomonas species in microbial fuel cells and biosurfactant production. ChemSusChem. 2024;17(4):e202401324.
- Palleroni NJ. The Pseudomonas story. Environ Microbiol. 2010;12(6):1377–1383.