Pseudomonas putida: Versatile Metabolic Hub for Environmental and Industrial Sustainability
Overview of the Microbe Pseudomonas putida KT2440 is a non‑spore‑forming, motile, Gram‑negative bacterium isolated from soil and rhizosphere environments[1]. It harbors a ~6.18 Mbp circular chromosome...
- Overview of the Microbe
- Bioremediation of Organic Pollutants
- Phytohormone Production and Plant Growth Promotion
- Industrial Biocatalysis and Synthetic Biology Chassis
- Plant Growth Promotion and Stress Alleviation
- Sustainable Biopolymer Production
- Challenges and Future Potential
- Spotlight on Research: Pseudomonas putida KT2440 Chassis Domestication
- Conclusion
- References
- 1
How Pseudomonas putida was genetically modified
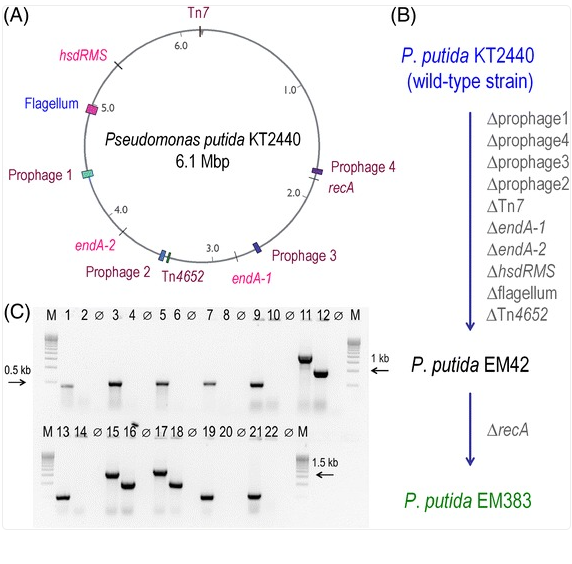
This figure shows how scientists edited the genome of Pseudomonas putida step by step, removing specific genes (like those for viruses and restriction systems) to create cleaner, more stable strains (EM42 and EM383).
- 2
Growth comparison between wild and engineered P. putida
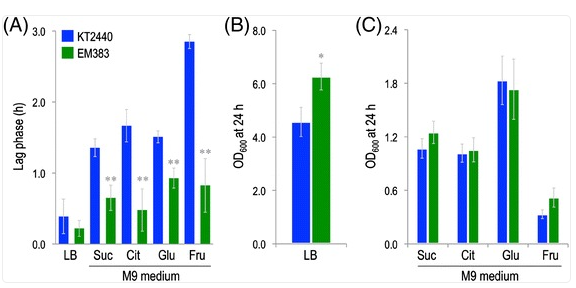
This figure compares how well the original strain (KT2440) and the engineered strain (EM383) grow on different sugar-based media.
Overview Table of Pseudomonas putida
- Feature
Description
- Scientific Name
Pseudomonas putida (Wells et al. 1981)
- Classification
Gram-negative rod; Phylum Proteobacteria; Class Gammaproteobacteria
- Habitat
Soil, freshwater, rhizosphere, polluted environments
- Key Functions
Aromatic‐compound degradation, biocatalysis, plant–microbe interactions
- Notable Abilities
Versatile metabolism, biofilm formation, solvent tolerance
- Applications
Bioremediation, industrial biocatalysis, bio-based polymer synthesis
- Genetic Engineering Potential
Broad host-range plasmids; CRISPR/Cas tools; genome‐reduced strains
- Challenges
Regulatory approval for release; metabolic burden in engineered strains
- Future Prospects
AI-guided pathway design; integrated bioprocessing of waste streams
Overview of the Microbe#
Pseudomonas putida KT2440 is a non‑spore‑forming, motile, Gram‑negative bacterium isolated from soil and rhizosphere environments[1]. It harbors a ~6.18 Mbp circular chromosome with over 5 600 protein‑coding genes, including numerous transporters, oxygenases, and regulatory systems that empower growth on diverse carbon sources and survival under physicochemical stresses[1]. A spontaneous restriction‑deficient derivative of strain mt‑2 (pWW0‑cured), KT2440 was certified as a Host–Vector Safety Level 1 system by the NIH in 1981, confirming its containment properties and non‑pathogenic status. Comparative genomics and metabolic modeling have further highlighted its robustness as a biosafety chassis for gene cloning and biotechnological exploitation[4].

Bioremediation of Organic Pollutants#
KT2440 degrades toluene, xylene, benzoate, and related mono‑ and polycyclic aromatics via chromosomal catabolic pathways and via the TOL plasmid pWW0, which carries xyl genes within transposons Tn4651 and Tn4653[1]. These pathways employ mono‑ and dioxygenases to convert recalcitrant compounds into Krebs‑cycle intermediates, enabling complete mineralization[2].
Phytohormone Production and Plant Growth Promotion#
Aromatic Hydrocarbon Degradation#
KT2440 degrades toluene, xylene, benzoate, and related mono‑ and polycyclic aromatics via chromosomal catabolic pathways and via the TOL plasmid pWW0, which carries xyl genes within transposons Tn4651 and Tn4653[1]. These pathways employ mono‑ and dioxygenases to convert recalcitrant compounds into Krebs‑cycle intermediates, enabling complete mineralization[2].
Xenobiotic and Emerging Contaminant Removal#
Beyond classic pollutants, P. putida strains transform styrene into biodegradable polyhydroxyalkanoate, offering an eco‑friendly route to valorize toxic waste. Chromosomal insertion of catabolic gene cassettes from other IncP plasmids (e.g., pCAR1) has extended KT2440’s substrate range to carbazole and chlorinated aromatics[3].
Industrial Biocatalysis and Synthetic Biology Chassis#
P. putida KT2440 combines high solvent tolerance—mediated by RND efflux pumps—and robust redox balance, making it an attractive host for biocatalysis under harsh industrial conditions[5]. A suite of modular vectors (SEVA plasmids), chromosomal “landing pads,” and CRISPR/Cas‑based editors enable stable expression of heterologous pathways and genome refactoring[9][8]. Recent reviews chart the domestication of KT2440 from a pollutant‑degrader to a standard synthetic‑biology chassis, highlighting incremental enhancements in genome accessibility, regulatory circuits, and metabolic predictability[7][5].
Plant Growth Promotion and Stress Alleviation#
Root colonization by KT2440 elicits induced systemic resistance (ISR) in maize, mediated by jasmonate‑ and ethylene‑dependent signaling pathways, leading to enhanced protection against fungal and bacterial pathogens[6]. Siderophore production (pyoverdine) chelates Fe³⁺ under iron‑limiting soils, increasing iron bioavailability for plants and inhibiting phytopathogen growth via iron competition[6]. These traits position P. putida as a bioinoculant for sustainable agriculture.
Sustainable Biopolymer Production#
KT2440 naturally accumulates medium‑chain‑length polyhydroxyalkanoates (mcl‑PHAs) up to ~40 % cell dry weight when cultured on fatty acids or lignin‑derived aromatics, offering a route to biodegradable plastics[11]. Quantum‑mechanics‑informed modeling of C/N ratios has optimized mcl‑PHA yields to ~13.8 % CDM at 40:1, demonstrating how in silico frameworks can guide fermentation strategies[10]. Genome‑scale metabolic reconstructions (iJN746) further aid in identifying metabolic bottlenecks and engineering high‑yield PHA producers[4].
Challenges and Future Potential#
Despite its advantages, KT2440 faces challenges in scaling—from biocontainment of engineered strains to streamlining regulatory approval across jurisdictions. Expanding the repertoire of characterized landing pads for heterologous genes will improve predictability and safety. Advances in dynamic sensor–regulator systems allow real‑time metabolic control, while multi‑omics and machine learning promise to accelerate chassis optimization[12].
Spotlight on Research: Pseudomonas putida KT2440 Chassis Domestication#
Brief Overview#
de Lorenzo et al. (2024) review the incremental engineering steps that transformed KT2440 from an environmental degrader to a premier synthetic‑biology chassis, detailing plasmid curing, genome editing tools, and standardized vector architectures[7].
Key Insights#
They document the removal of TOL pWW0, the adoption of SEVA plasmids, the mapping of safe genomic insertion loci, and the integration of CRISPR/Cas and lambda‑Red tools, creating a highly tractable host for modular pathway assembly[7].
Why This Matters#
By codifying a “domestication roadmap,” this work enables reproducible chassis deployment across laboratories and industries, lowering barriers to innovative bioprocess development.
Summary Table: Spotlight Study#
| Category | Details |
| Lead Researchers | Victor de Lorenzo; Danilo Pérez‑Pantoja; Pablo I. Nikel |
| Affiliations | Systems Biology Dept., CNB‑CSIC, Madrid, Spain; Universidad Tecnológica Metropolitana, Santiago, Chile; Novo Nordisk Foundation Center for Biosustainability, Denmark |
| Research Focus | Chassis domestication for synthetic biology |
| Key Breakthroughs | TOL plasmid curing; standardized SEVA vector adoption; genome editing (CRISPR/Cas, lambda‑Red); identification of safe genomic “landing pads” for heterologous genes |
| Collaborative Efforts | CNB‑CSIC (Spain), UTEM & CAPES (Chile), Novo Nordisk Center (Denmark) |
| Published Work | Journal of Bacteriology |
| Publication Date | July 2024 |
| Location | Haryana (India) |
| Key Findings | Established a unified toolkit and genome modifications that underpin KT2440’s widespread use as a robust, modular chassis in metabolic engineering and biocatalysis[7]. |
Conclusion#
Pseudomonas putida KT2440 epitomizes a versatile metabolic hub whose natural biodegradation capacities have been harnessed and refined through synthetic‑biology tools to address environmental pollution, sustainable agriculture, and green polymer production. Genome‑scale models, dynamic regulation systems, and standardized genetic toolkits continue to expand its utility, while ongoing work in biocontainment and chassis optimization will ensure safe, scalable deployment in industrial and agricultural settings.
References#
- Nelson KE, Weinel C, Paulsen IT, Dodson RJ, Hilbert H, Martins dos Santos VAP, et al. Complete genome sequence and comparative analysis of the metabolically versatile Pseudomonas putida KT2440. Environ Microbiol. 2002;4(12):799–808. doi:10.1046/j.1462-2920.2002.00304.x. PubMed
- Puchałka, J., Oberhardt, M. A., Godinho, M., Bielecka, A., Regenhardt, D., Timmis, K. N., Papin, J. A., & Martins dos Santos, V. A. P. (2008). Genome-scale reconstruction and analysis of the Pseudomonas putida KT2440 metabolic network facilitates applications in biotechnology. PLoS Computational Biology, 4(10), e1000210. https://doi.org/10.1371/journal.pcbi.1000210
- Miyakoshi, M., Shintani, M., Terabayashi, T., Kai, S., Yamane, H., & Nojiri, H. (2007). Transcriptome analysis of Pseudomonas putida KT2440 harboring the completely sequenced IncP-7 plasmid pCAR1. Journal of Bacteriology, 189(19), 6585–6594. https://doi.org/10.1128/JB.00684-07
- Sonnenschein EC, et al. A genome‑scale reconstruction of Pseudomonas putida KT2440’s metabolism. BMC Syst Biol. 2008;2:79. doi:10.1186/1752-0509-2-79. BioMed Central
- Nikel PI, Martínez‑García E, de Lorenzo V. Biotechnological domestication of pseudomonads using synthetic biology. Nat Rev Microbiol. 2014;12:368–379. doi:10.1038/nrmicro3253. Nature
- Planchamp C, Glauser G, Mauch‑Mani B. Root inoculation with Pseudomonas putida KT2440 induces transcriptional and metabolic changes and systemic resistance in maize plants. Front Plant Sci. 2014;5:719. doi:10.3389/fpls.2014.00719. Nature
- de Lorenzo V, Pérez‑Pantoja D, Nikel PI. Pseudomonas putida KT2440: the long journey of a soil‑dweller to become a synthetic biology chassis. J Bacteriol. 2024;206(7):e0013624. doi:10.1128/jb.00136-24. PubMed
- Martínez‑García E, de Lorenzo V. Molecular tools and emerging strategies for deep genetic/genomic refactoring of Pseudomonas. Curr Opin Biotechnol. 2017;47:120–132. doi:10.1016/j.copbio.2017.06.013. ScienceDirect
- Chaves, J. E., Wilton, R., Gao, Y., Muñoz Muñoz, N., Burnet, M. C., Schmitz, Z., Rowan, J., Burdick, L. H., Elmore, J., Guss, A., Close, D., Magnuson, J. K., Burnum-Johnson, K. E., & Michener, J. K. (2020). Evaluation of chromosomal insertion loci in the Pseudomonas putida KT2440 genome for predictable biosystems design. Metabolic Engineering Communications, 11, e00139. https://doi.org/10.1016/j.mec.2020.e00139
- Ho LYL, Pan L, Meng F, Ho KTM, Liu F, Wu MT, et al. Quantum modeling simulates nutrient effect of bioplastic polyhydroxyalkanoate (PHA) production in Pseudomonas putida. Sci Rep. 2024;14:18255. doi:10.1038/s41598-024-68727-7. Nature
- Huijberts GNM, Eggink G. Production of poly(3‑hydroxyalkanoates) by Pseudomonas putida KT2442 in continuous cultures. Appl Microbiol Biotechnol. 1996;46(3):233–239. doi:10.1007/s002530050810. PMC
- Wang Z, Nomura CT. Synthetic control of metabolic states in Pseudomonas putida by dynamic sensor–regulator integration. mBio. 2021;12(1):e01794-21. doi:10.1128/mBio.01794-21. ASM Journals