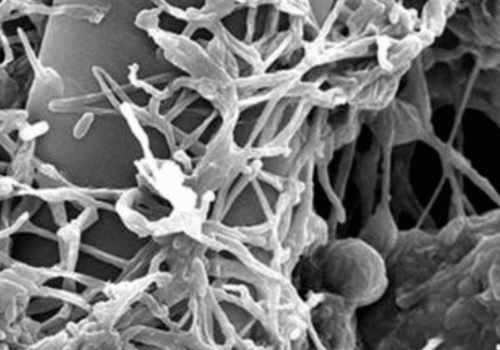
Coniothyrium minitans: A Mycoparasitic Ally for Sustainable Crop Protection
Overview of the Microbe Coniothyrium minitans, also known by the synonym Paraphaeosphaeria minitans, is an ascomycete fungus in the family Didymosphaeriaceae [1]. This obligate mycoparasite...
- Overview of the Microbe
- Biocontrol of Sclerotinia Diseases
- Mycoparasitism Mechanisms and Enzyme Secretion
- Soil Health Enhancement and Plant Growth Promotion
- Formulation Strategies and Field Applications
- Challenges and Future Potential
- Spotlight on Research: Whole‑Genome Sequencing of Coniothyrium minitans
- Conclusion
- References
- 1
Protein Functions in the Genome of C. minitans
This figure shows how the proteins made by C. minitans are grouped based on their roles, helping scientists understand what the fungus is capable of.
- 2
Gene Activity Changes in C. minitans When Exposed to Another Fungus
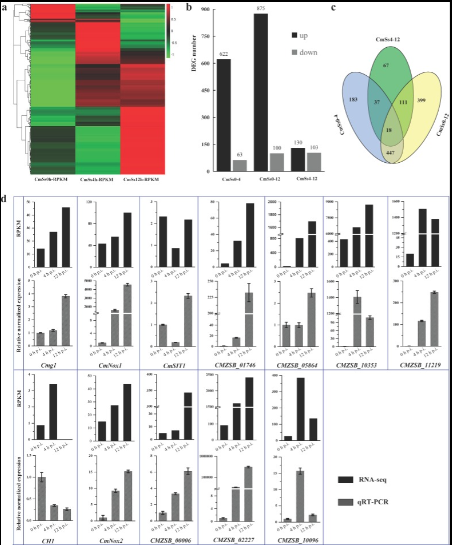
This figure shows which genes in C. minitans become more or less active when it encounters the fungus S. sclerotiorum, and confirms these changes using a gene-testing method
Overview Table of Coniothyrium minitans
- Feature
Description
- Scientific Name
Coniothyrium minitans
- Classification
Ascomycete fungus; Phylum Ascomycota; Order Pleosporales
- Habitat
Soil, plant debris, rhizosphere
- Key Functions
Mycoparasitism of plant-pathogenic fungi; organic matter decomposition
- Notable Abilities
Production of chitinases, glucanases; sclerotial degradation; spore resilience
- Applications
Biocontrol of Sclerotinia spp.; integrated pest management; soil amendment
- Genetic Engineering Potential
Candidate for enhancement of lytic enzyme genes; marker-assisted strain improvement
- Challenges
Consistent performance under varied field conditions; formulation shelf-life; regulation
- Future Prospects
Genomic-guided strain selection; synergistic consortia; integration with circular-bioeconomy
Overview of the Microbe#
Coniothyrium minitans, also known by the synonym Paraphaeosphaeria minitans, is an ascomycete fungus in the family Didymosphaeriaceae [1]. This obligate mycoparasite targets sclerotia; hardened survival structures of plant-pathogenic fungi, most notably Sclerotinia sclerotiorum, S. minor, and Sclerotium cepivorum. Its global presence is strongly correlated with regions where these pathogens occur, and it is found across temperate agricultural and natural soils [2].
Microscopically, C. minitans produces darkly pigmented, septate hyphae capable of penetrating the sclerotial cortex and colonizing internal tissues [3]. It reproduces asexually via ovoid to ellipsoid conidia formed within black pycnidia that rupture to disseminate spores. Laboratory studies show spore germination in moist soil at temperatures between 10 °C and 25 °C, peaking around 20 °C and declining sharply above 30 °C [4].
Ecological assessments indicate C. minitans cannot survive as a free-living saprophyte. Instead, its spores and mycelia are strictly associated with sclerotia, and overall soil populations decline in their absence. However, infected sclerotia can preserve viable spores and pycnidia for up to 18 months under favorable conditions [5].

Biocontrol of Sclerotinia Diseases#
Biological control using Coniothyrium minitans centers on its capacity to suppress Sclerotinia sclerotiorum and S. minor through targeted attack on their sclerotia. When applied to soil three months before expected disease onset, C. minitans can reduce sclerotia viability by up to 95% and decrease apothecial (fruiting body) production by approximately 90%, resulting in 10–70% reductions in disease symptoms [6]. Field trials in canola and soybean demonstrate that soil incorporation of C. minitans leads to fewer sclerotia and improved yield stability [6, 7].Comparative glasshouse studies further show that C. minitans performs comparably or superiorly, to fungicides when applied in early crop stages, particularly against lettuce drop and oilseed rape stem rot [6, 8]. Notably, integration with Bacillus amyloliquefaciens has enhanced soybean white mold control, suggesting synergistic outcomes within microbial consortia [9].
Mycoparasitism Mechanisms and Enzyme Secretion#
The mycoparasitism of Coniothyrium minitans against Sclerotinia sclerotiorum is orchestrated through a coordinated deployment of hydrolytic enzymes, signaling pathways, detoxification systems, and antifungal metabolites. Transcriptome profiling of strain ZS-1 during early host interaction revealed the upregulation of 17 genes encoding fungal cell-wall–degrading enzymes, especially chitinases and β‑1,3‑glucanases, essential for breaching host structures [2]. Among these, the cmg1 β‑1,3‑glucanase and CH1 chitinase genes show dramatic increases, consistent with enhanced extracellular enzymatic activity [2].
Concurrent activation of key regulatory systems—namely the NADPH oxidase complex (CmNox1) and MAP‑kinase cascade via CmSlt2—is critical; disruption of CmNox1 results in loss of both conidiation and parasitic ability [10]. Additionally, a pH-responsive transcription factor (CmpacC) modulates chitinase and glucanase expression while repressing Cmoxdc1, the gene encoding oxalate decarboxylase [11]. Oxalate decarboxylase is vital for neutralizing oxalic acid—a pathogenicity factor produced by Sclerotinia—thereby enabling colonization under acidic conditions.
Complementing these molecular investments, C. minitans synthesizes antifungal metabolites such as macrosphelide A, and expresses transporter proteins, heat-shock factors, peroxisomal proteins, and proteolytic enzymes that further enhance its parasitic competency [2, 12].
Soil Health Enhancement and Plant Growth Promotion#
By reducing viable Sclerotinia sclerotia in soil, Coniothyrium minitans plays a key role in improving soil health and indirectly supporting plant growth. Field studies report that populations of C. minitans can persist in non-irrigated soils for up to 750 days, declining from ~7×10⁵ to ~4×10⁴ cfu/g of soil, indicating long-term colonization when sclerotia are present [12]. This persistence parallels sustained reductions in sclerotial viability and apothecial production, which in turn minimizes disease inoculum affecting subsequent crops [6].
Environmental research confirms that C. minitans thrives under moderate moisture (6–45%) and temperature (4–28 °C) regimes, but declines under extreme heat (>45 °C) [12]. Importantly, regulatory data show that application of commercial products, such as Contans® WG, does not adversely affect non-target soil microbiota or mesofauna, including earthworms and collembolans; it does not leach or persist in soil beyond target zones [13, 14]
Although C. minitans is not a direct plant growth promoter, its pathogen suppression function supports root health and enhances nutrient access, leading to healthier plants and stabilized yields over rotations [6].
Formulation Strategies and Field Applications#
Effective formulation and application of Coniothyrium minitans hinge on delivering viable spores where sclerotia are present and supporting fungal growth under field conditions. The most widely used commercial product, Contans® WG (wettable granules), contains approximately 1.0 × 10¹² CFU/kg, equivalent to 50 g spores per kilogram of formulation [16]. Regulatory labels advise soil application 3 to 4 months before disease onset (or planting), followed by incorporation into the top 2 inches, optimizing contact between spores and sclerotia [14].
Field experiments in peanut rotations demonstrated that applying Contans® WG at 2 kg/ha annually over three years significantly reduced Sclerotinia blight incidence and sclerotial counts, with similar outcomes at both 2 kg/ha and 4 kg/ha rates [17]. Trials in oilseed rape, using a maize-perlite granule with C. minitans, showed 80–90% suppression of apothecia and disease compared to untreated controls. Contans® WG is effective across varied crops—including carrot, lettuce, canola, sunflower, and beans—and is approved for organic use in some regions [18].
Challenges and Future Potential#
Despite its success, Coniothyrium minitans faces constraints under variable field conditions. Conidial survival and infection efficacy decline sharply in environments with high rainfall or intense irrigation, likely due to wash-off from the soil or plant surfaces [19]. Its biocontrol activity also diminishes when temperatures exceed 28–30 °C or soil moisture remains below 15%, limiting its effectiveness in hot or arid regions [14].
Ensuring consistent strain performance remains a challenge. According to EFSA 2016, [14] post-application Coniothyrium minitans populations typically decline by two orders of magnitude within the first week, then stabilize for up to six months. However, viability and infection capacity drop sharply at soil temperatures above 25 °C. Additionally, shelf-life studies from the Contans® WG technical dossier [16], show a 25% reduction in spore activity after 18 months, underscoring the need for robust strains and optimized formulation for commercial consistency.
Nevertheless, advanced formulation approaches such as encapsulating pycnidia in alginate-cellulose matrices have shown enhanced survival at both low (4 °C) and moderate (28 °C) storage temperatures [20]. The discovery and characterization of additional oxalate-degrading genes, such as CmOxdc3, provide molecular targets to engineer strains with improved pH tolerance and parasitic efficacy [15].
Spotlight on Research: Whole‑Genome Sequencing of Coniothyrium minitans#
Brief Overview#
In January 2021, Patel et al. [21] reported the first draft genome sequence of C. minitans strain Conio (IMI 134523), the strain used in commercial formulations such as Contans® WG (Patel et al., 2021, Mol. Plant-Microbe Interact.). The 47.9 Mb assembly comprises 34 scaffolds with 48.06% GC content, achieved at approximately 244× coverage and with 97.7% completeness based on Ascomycota BUSCO analysis [21].
Key Insights#
The genome contains approximately 13,677 predicted genes, including 34 bZIP transcription factors, numerous carbohydrate-active enzymes (CAZymes), secreted hydrolases, and 15 secondary metabolite biosynthesis clusters, including genes for epipolythiodioxopiperazine (ETP)-type metabolites [21]. This indicates a strong parasitic arsenal directed at sclerotial cell walls, detoxification, and host suppression.
Why This Matters#
This genomic resource enables precise functional analyses, strain improvement, and molecular marker development for quality control. Identification of genes contributing to stress resilience, parasitism, and secondary metabolite production can guide next-generation biocontrol formulations.
Summary Table: Spotlight Study#
| Category | Details |
| Lead Researchers | Patel D., Shittu T.A., Baroncelli R., Muthumeenakshi S., Osborne T.H., Janganan T.K., Sreenivasaprasad S. |
| Affiliations | University of Bedfordshire (UK); Universidad de Salamanca (Spain) |
| Research Focus | Draft genome assembly and annotation of strain Conio (IMI 134523) |
| Key Breakthroughs | 47.9 Mb, 34 scaffolds, 97.7% BUSCO completeness; 13,677 genes; 34 bZIP TFs; 15 secondary metabolite clusters |
| Collaborative Efforts | Academic–industry collaboration enabling product registratio |
| Published Work | “Genome Sequence of the Biocontrol Agent Coniothyrium minitans Conio (IMI 134523)” |
| Publication Date | February 2021 (published online January 18) |
| Location | UK & Spain |
| Key Findings | Comprehensive genome resource with parasitism and biosynthesis gene inventory |
Conclusion#
The molecular insights afforded by the 47.9 Mb draft genome of Coniothyrium minitans [21] reinforce its status as a precise and safe mycoparasite targeting Sclerotinia sclerotia. Coupling robust enzymatic, metabolic, and regulatory systems with advanced formulation strategies underscores its potential in sustainable agriculture. Emerging genomics-driven innovations, including strain enhancement and microbial consortia, promise to broaden its applicability and reliability in crop protection.
References#
- Verkley, G. J. M., da Silva, M., Wicklow, D. T., & Crous, P. W. (2004). Paraconiothyrium, a new genus to accommodate C. minitans and related anamorphs. Studies in Mycology, 50, 323–335.
- Zhao, H., Zhou, T., Xie, J., Cheng, J., Jiang, D., & Fu, Y. (2020). Mycoparasitism illuminated by genome and transcriptome sequencing of Coniothyrium minitans, an important biocontrol fungus of S. sclerotiorum. Microbial Genomics, 6(3), e000345. https://doi.org/10.1099/mgen.0.000345
- Ren, L., Li, G., Han, Y. C., Jiang, D. H., & Huang, H. C. (2007). Degradation of oxalic acid by Coniothyrium minitans and its effects on production and activity of β-1, 3-glucanase of this mycoparasite. Biological Control, 43(1), 1-11. https://doi.org/10.1016/j.biocontrol.2007.06.006
- Sandys‑Winsch, C., Whipps, J. M., Gerlagh, M., & Kruse, M. (1993). World distribution of the sclerotial mycoparasite Coniothyrium minitans. Mycological Research, 97, 1175–1178. https://doi.org/10.1016/S0953-7562(09)81280-X
- de Vrije, T., Antoine, N., Buitelaar, R. M., Bruckner, S., Dissevelt, M., Durand, A., … Whipps, J. M. (2001). The fungal biocontrol agent Coniothyrium minitans: production by solid-state fermentation, application and marketing. Applied Microbiology & Biotechnology, 56(1–2), 58–68. https://doi.org/10.1007/s002530100678
- Crop Protection Network. (2025). An overview of white mold. Retrieved July 2025, from https://cropprotectionnetwork.org/publications/an-overview-of-white-mold
- McLaren, D., Conner, R., & McAndrew, D. (2006). Impact of timing, rate and application technology on biological control of Sclerotinia stem rot of canola caused by Sclerotinia sclerotiorum (Final report CARP 2006‑03). Canola Research Hub. Retrieved July 2025, from https://www.canolacouncil.org/research-hub/impact-of-timing-rate-and-application-technology-on-biological-control-of-sclerotinia-stem-rot-of-canola-caused-by-sclerotinia-sclerotiorum/
- Budge, S. P., & Whipps, J. M. (2001). Potential for integrated control of Sclerotinia sclerotiorum in glasshouse lettuce using Coniothyrium minitans and reduced fungicide application. Phytopathology, 91(2), 221-227. https://doi.org/10.1094/PHYTO.2001.91.2.221
- Conrad, A. M., & Telenko, D. E. P. (2023). Efficacy of biocontrol agents Coniothyrium minitans and Bacillus amyloliquefaciens for managing Sclerotinia sclerotiorum in Indiana soybean. PhytoFrontiers, 3(3). https://doi.org/10.1094/PHYTOFR-07-22-0080-R
- Wei, W., Zhu, W., Cheng, J., Xie, J., Jiang, D., … & Fu, Y. (2016). Nox complex signal and MAPK cascade pathway are cross-linked and essential for pathogenicity and conidiation of mycoparasite Coniothyrium minitans. Scientific Reports, 6, 24325. https://doi.org/10.1038/srep24325
- Lou, Y., Han, Y.-C., Yang, L., Wu, M.-D., Zhang, J., Cheng, J.-S., & Jiang, D.-H. (2015). CmpacC regulates mycoparasitism, oxalate degradation and antifungal activity in the mycoparasitic fungus Coniothyrium minitans. Environmental Microbiology, 17(11), 4711–4729. https://doi.org/10.1111/1462-2920.13018
- Yang, L., Li, G. Q., Long, Y. Q., Hong, G. P., Jiang, D. H., & Huang, H. C. (2010). Effects of soil temperature and moisture on survival of Coniothyrium minitans conidia in central China. Biological Control, 55(1), 27–33. https://doi.org/10.1016/j.biocontrol.2010.06.010
- EPA. (2001). Biopesticide Registration Action Document: Coniothyrium minitans strain CON/M/91‑08. U.S. Environmental Protection Agency. https://www3.epa.gov/pesticides/chem_search/reg_actions/registration/decision_PC-028836_1-Aug-01.pdf
- EFSA. (2016). Peer review of the pesticide risk assessment of the active substance Coniothyrium minitans. EFSA Journal, 14(6), 4517. https://doi.org/10.2903/j.efsa.2016.4517
- Xu, Y., Wu, M., Zhang, J., Li, G., & Yang, L. (2022). Cloning and molecular characterization of CmOxdc3 coding for oxalate decarboxylase in the mycoparasite Coniothyrium minitans. Journal of Fungi, 8(12), 1304. https://doi.org/10.3390/jof8121304
- Bayer CropScience. (2005). Contans® WG technical dossier (pp. 1–13). Bayer AG. https://www.bayer.com/sites/default/files/M-465794-04-3_1.PDF
- Partridge, D. E., Sutton, T. B., Jordan, D. L., Curtis, V. L., & Bailey, J. E. (2006). Management of Sclerotinia blight of peanut with the biological control agent Coniothyrium minitans. Plant Disease, 90(7), 957–963. https://doi.org/10.1094/PD-90-0957
- Lallemand Plant Care. (2025). LALSTOP Contans® WG technical sheet. Retrieved from https://www.lallemandplantcare.com/en/usa/products/product-details/lalstop-contans-wg/
- Green, I., & Whipps, J. M. (1998). Splash dispersal of Coniothyrium minitans in the glasshouse. Annals of Applied Biology, 133(1), 45–53. https://doi.org/10.1111/j.1744-7348.1998.tb05186.x
- Moretini, A., & Melo, I. S. D. (2007). Formulation of Coniothyrium minitans for biocontrol of white mold caused by Sclerotinia sclerotiorum. Pesquisa Agropecuária Brasileira, 42, 155-161. https://doi.org/10.1590/S0100-204X2007000200003
- Patel, D., Shittu, T. A., Baroncelli, R., Muthumeenakshi, S., Osborne, T. H., Janganan, T. K., & Sreenivasaprasad, S. (2021). Genome sequence of the biocontrol agent Coniothyrium minitans conio (IMI 134523). Molecular Plant-Microbe Interactions, 34(2), 222-225. https://doi.org/10.1094/MPMI-05-20-0124-A