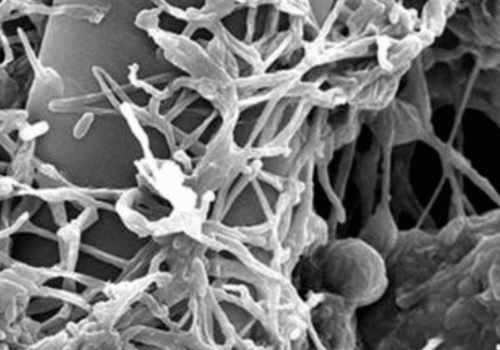
Verticillium lecanii: A White Muscardine Entomopathogen and Biocontrol Agent
Overview of the Microbe Verticillium lecanii (Zimmermann) Viegas is an entomopathogenic fungus historically known for causing “white muscardine” disease in insects. It is now classified...

- Overview of the Microbe
- Entomopathogenic Mechanisms and Pest Management
- Mass Production, Formulation and Field Efficacy
- Chitinase Secretion and Secondary Metabolites
- Plant Endophytism and Growth Promotion
- Taxonomy, Genetic Tools and Strain Improvement
- Challenges and Future Potential
- Spotlight on Research: Verticillium lecanii Elicitor for Pest Control
- Conclusion
- References
- 1
Shows that the insect population dropped more when feeding on PeVL1-treated plants compared to untreated ones.
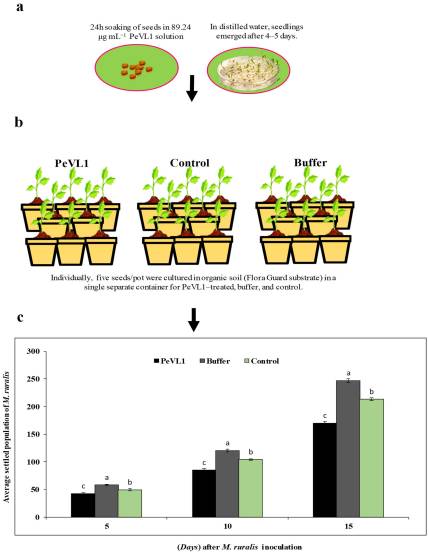
PeVL1 treatment reduced insect population more than the control.
- 2
Reveals that insects feeding on PeVL1-treated plants had fewer offspring, and all generations grew more slowly.
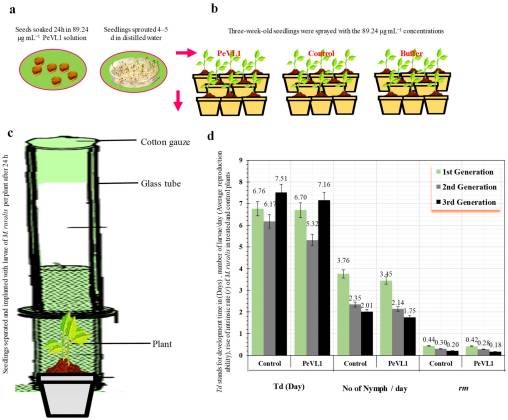
PeVL1 lowered insect reproduction and slowed growth across generations.
Overview Table of Verticillium lecanii
- Feature
Description
- Scientific Name
Verticillium lecanii (Zimmerman) Viegas 1939
- Classification
Kingdom: Fungi; Phylum: Ascomycota; Class: Sordariomycetes; Order: Hypocreales; Family: Cordycipitaceae (Wikipedia)
- Habitat
Soil, phyllosphere, insect cadavers; tropical and temperate regions
- Key Functions
Entomopathogenesis; chitinase and protease secretion; endophytic colonisation
- Notable Abilities
Rapid cuticle penetration; broad host range (Hemiptera, Homoptera, Thysanoptera, Coleoptera)
- Applications
Biopesticides against aphids, whiteflies, thrips; plant bioinoculants
- Genetic Engineering Potential
Agrobacterium-mediated transformation; protoplast PEG methods
- Challenges
Variable field efficacy; UV and desiccation sensitivity; regulatory harmonisation
- Future Prospects
Formulation improvements; endophytic strain development; CRISPR-based virulence tuning
Overview of the Microbe#
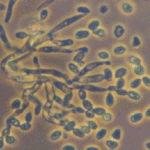
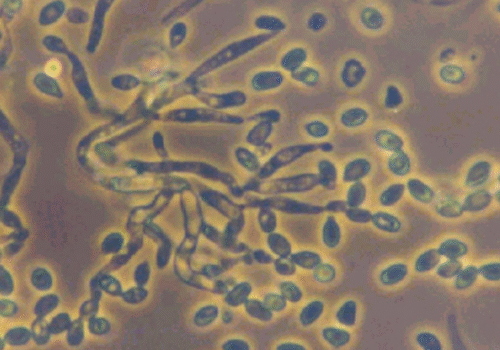
Verticillium lecanii (Zimmermann) Viegas is an entomopathogenic fungus historically known for causing “white muscardine” disease in insects. It is now classified as Lecanicillium lecanii in the family Cordycipitaceae (order Hypocreales), reflecting a taxonomic reorganization based on genetic studies. Many isolates once identified as V. lecaniihave been reclassified into related species (e.g. Lecanicillium muscarium, L. longisporum, L. attenuatum) due to improved molecular differentiation. L. lecanii itself is primarily a pathogen of soft-bodied homopteran insects such as scale insects (Coccidae), but the broader Lecanicillium group targets a wide range of pests including aphids, whiteflies, thrips, and mealybugs.
This fungus is ubiquitous in soils and humid habitats worldwide and was first noted as an insect pathogen in the 19th century (initially described on scale insects in 1898 by Zimmerman as Cephalosporium lecanii). Under conducive conditions, V. lecanii infects insect hosts upon contact, ultimately killing them and producing an outward eruption of white fuzzy mycelium and spores (hence the “white halo” or muscardine appearance). It has been developed as a biocontrol agent for agricultural and horticultural use, especially in greenhouse crops. In the 1970s, researchers R.A. Hall and H.D. Burges at the Glasshouse Crops Research Institute (UK) pioneered the use of V. lecanii to control pest populations of aphids, whiteflies, and thrips in glasshouses. Commercial biopesticide products derived from Lecanicillium fungi include Mycotal (based on L. muscarium) and Vertalec (based on L. longisporum), distributed by Koppert Biological Systems, among others. These products and other formulations are applied as spore suspensions to foliage, where the fungus can infect target pests. V. lecanii is valued in integrated pest management for its host specificity and environmental safety, posing minimal risk to plants and beneficial insects when used correctly.

Entomopathogenic Mechanisms and Pest Management#
Like many entomopathogenic fungi, Verticillium lecanii infects insects through a combination of contact adhesion, enzymatic penetration, and internal proliferation. Infection begins when a sticky conidium (spore) attaches to the host insect’s cuticle. The spore germinates and forms a specialized structure called an appressorium that exerts mechanical pressure on the cuticle. Simultaneously, the fungus secretes cuticle-degrading enzymes – notably proteases, lipases, and chitinases – to breach the insect’s exoskeleton. Once through the cuticle, the fungal hyphae invade the hemocoel (body cavity), where they proliferate as yeasty blastospores, consuming the insect’s internal tissues and releasing toxins that contribute to host death. The infected insect usually succumbs within several days as its organs are destroyed[2]. After the host’s death, V. lecanii often produces external white mycelium and spores on the cadaver (especially under humid conditions), which can spread to new hosts (hence the visibly white “muscardine” mold on corpses, as seen in the figure above).
V. lecanii has a broad host range among sap-sucking insect pests and is deployed for biocontrol of aphids, whiteflies, thrips, mealybugs, scale insects, and others. Notably, it was shown to effectively control greenhouse aphids and whiteflies in early trials. Field and greenhouse studies have demonstrated that sprays of V. lecanii spore suspensions can infect and kill significant portions of pest populations under optimal conditions. For example, at 25 °C and near 100% relative humidity, V. lecanii can achieve nearly 100% mortality of certain aphid species within ~2–3 weeks. Typically, pests die 4–7 days after infection, and outbreaks can be suppressed when the fungus is applied preventatively or at early infestation stages. An advantage of V. lecanii in pest management is the contact mode of action – insects do not need to ingest spores (as the fungus penetrates directly through the cuticle), making it effective against piercing-sucking insects that are often hard to target with poisons. Furthermore, V. lecanii and related species rarely engender resistance in pest populations, in contrast to chemical insecticides, because the fungi attack via can co-evolve with hosts. These fungi are considered environmentally friendly and can be used in rotation or combination with other biocontrol agents. Studies have shown Lecanicillium species promultiple mechanisms andvide significant control of aphids, whiteflies, thrips, and even certain leafhoppers[1]. Consequently, V. lecanii has become a key tool in integrated pest management (IPM), especially in protected cultivation systems like greenhouses where maintaining the high humidity needed for fungal infection is feasible.
Mass Production, Formulation and Field Efficacy#
Effective use of V. lecanii as a biocontrol agent requires reliable mass-production of infective propagules (spores) and formulation into stable products. Mass production of Lecanicillium fungi is achieved either by solid-state fermentation on grains or other substrates, or by liquid fermentation to produce high spore yields. For instance, culturing V. lecanii on cooked rice or rice bran can produce on the order of 1–2 × 10^9 conidia per gram of substrate. Industrial producers often use a biphasic production system: the fungus is first grown in liquid media (to generate mycelial biomass or blastospores), then transferred to a nutritive solid substrate to stimulate abundant aerial conidia formation. This two-stage method improves yield and reduces contamination risk, resulting in highly concentrated spore powders. V. lecanii spores are typically formulated as wettable powders (WP) or oil dispersions that can be rehydrated and sprayed. Key formulation considerations include spore viability and shelf-life – spores must remain alive but dormant in the product. Research has shown that adding desiccants, nutritional adjuncts (like rice starch or whey), and sticking agents (like jaggery or molasses) to formulations can enhance spore survival and efficacy on crops.Field efficacy of V. lecanii biopesticides depends strongly on environmental conditions. High humidity (≥ 90% RH) is usually required for spores to germinate on the insect cuticle and infect the host. In fact, V. lecanii conidia typically only germinate in the presence of a water film or saturated humidity, making this fungus most effective in humid greenhouse environments or tropical climates. Temperature is another critical factor: optimal infection occurs around 20–28 °C, with diminished activity at cooler or hotter extremes. Field trials in tea plantations, for example, found L. lecanii gave effective thrips control when applied in the early morning or evening (to avoid UV exposure) under warm, humid conditions. Ultraviolet light and heat are detrimental to fungal spores – even 30 minutes of direct sun can significantly reduce V. lecanii spore viability on leaves. To mitigate this, formulations often include UV-protectants or the fungus is applied when solar radiation is low. Despite these challenges, V. lecanii has shown good field performance in protected and semi-protected crops. For instance, a wettable powder formulation of L. lecanii applied at ~10^7 conidia/mL was effective in suppressing tea thrips in field trials, without harming natural predator insects. Numerous Lecanicillium-based biocontrol products (over a dozen worldwide) have been registered for use on ornamentals, vegetables, field crops, and even stored products. When mass-produced and applied properly, V. lecanii provides a sustainable alternative to chemical pesticides, though maintaining ideal microclimatic conditions (humidity and temperature) is key to its success in the field.
Chitinase Secretion and Secondary Metabolites#
One of the notable virulence factors of Verticillium lecanii is its secretion of chitinases, enzymes that degrade chitin – a major component of the insect exoskeleton. During the infection process, V. lecanii produces multiple chitinase isoforms that work in concert with proteases to break down the host cuticle and facilitate fungal penetration. These chitinases are considered pathogenicity determinants: experimentally, overexpression of chitinase genes in related entomopathogens (e.g. Beauveria bassiana) led to increased virulence, indicating that higher chitinase activity accelerates host kill. In V. lecanii, at least two chitinase genes have been characterized. One example is Vlchit1, an endochitinase gene encoding a 423-amino-acid enzyme, which has been cloned and studied for its potential in strain improvement[2]. The Vlchit1 enzyme has a typical TIM barrel structure and actively hydrolyzes chitin, helping the fungus breach insect cuticles[2]. Such enzymes not only assist in infection but could be exploited in transgenic approaches (for instance, engineering plants or other microbes to express V. lecanii chitinases for pest resistance). The strong chitinolytic arsenal of V. lecanii underpins its ability to infect a wide range of hosts, especially those with protective exoskeletons or egg shells.
In addition to enzymatic action, V. lecanii produces several secondary metabolites that contribute to its entomopathogenic and antimicrobial activity. Notably, the fungus can synthesize bassianolide, a cyclodepsipeptide toxin also found in Beauveria bassiana. Bassianolide exhibits potent insecticidal properties; it was originally isolated in 1977 and is known to be lethal to various insects by disrupting their physiology. This compound is used as a benchmark bioactive in some fungal insecticide formulations. V. lecanii is also reported to produce other metabolites such as dipicolinic acid and unique compounds called vertilecanins, as well as various fatty acids and peptides, though these are less well characterized. Early research by N. Claydon et al. (1982) identified several insecticidal compounds in V. lecaniiculture filtrates. The combined action of these metabolites can weaken the host’s immune defenses and cause toxicosis, complementing the fungus’s direct tissue destruction. Interestingly, beyond its insect targets, V. lecanii exhibits antimicrobial effects – some strains secrete metabolites that inhibit plant pathogenic fungi (for example, substances that suppress rust and mildew fungi on plants). This broad biochemical repertoire makes V. lecanii a candidate for multitarget biocontrol: it can directly infect pests and potentially protect plants via antimicrobial compounds. Current research is exploring ways to enhance secondary metabolite production (e.g. through nutrient manipulation or genetic modification) to improve the efficacy of V. lecanii-based biopesticides.
Plant Endophytism and Growth Promotion#
Beyond its role as an insect pathogen, Verticillium lecanii can also behave as a beneficial endophyte and mycoparasitein plant systems. Studies have shown that V. lecanii (and related Lecanicillium species) are capable of colonizing plant tissues asymptomatically, which can confer advantages such as induced systemic resistance and protection against plant pathogens. For instance, research by Benhamou and Brodeur (2001) demonstrated that inoculating cucumber roots with V. lecanii could induce host defense reactions that protected the plants from Pythium ultimum root rot. The fungus acted as a mycoparasite and elicitor: it colonized the rhizosphere/roots and primed the plant’s immune system (e.g. by triggering production of defensive enzymes and structural barriers), thereby reducing disease severity. In practical terms, this means V. lecanii might serve a dual role – attacking insect pests and simultaneously suppressing certain soil-borne diseases. Similarly, V. lecanii has been observed parasitizing other fungi; for example, it can infect the spores and fruiting bodies of the chrysanthemum white rust fungus (Puccinia horiana), thereby acting as a hyperparasite that curbs rust outbreaks. This kind of antagonism suggests V. lecanii could be integrated into crop protection not only for insect control but also as a biological control for some fungal pathogens.
As an endophyte, Lecanicillium species may also promote plant growth and stress tolerance. Although specific plant growth-promoting traits of V. lecanii are still being studied, endophytic entomopathogenic fungi in general have been linked to enhanced nutrient uptake, growth hormone production, and improved tolerance to abiotic stresses in their host plants. There is evidence that V. lecanii can internally colonize certain crop plants (such as cotton and tomato) without causing disease, providing a kind of systemic protection against pests. For example, when L. lecanii grows within plant tissues, the plant may have faster induction of defense genes upon insect attack, resulting in reduced herbivore damage. One study found that endophytic colonization by L. lecanii in tomato led to decreased survival of aphids feeding on those plants, presumably because the fungus-produced metabolites or plant defense compounds negatively affected the insects (a form of bottom-up pest resistance). Additionally, the presence of Lecanicillium in plants can elicit the salicylic acid (SA) signaling pathway (commonly associated with systemic acquired resistance), which helps the plant fend off not just insects but also some pathogens. While the plant-growth-promotion effects of V. lecanii are less pronounced than those of some bacterial endophytes, the fungus can indirectly improve plant health by reducing pest pressure and disease incidence.
Overall, the ability of Verticillium lecanii to act as an endophyte and defensive symbiont expands its usefulness in agriculture. It hints at future applications where crops could be “inoculated” with the fungus (for example, seed treatments or root drenches) so that the plant enjoys ongoing protection from both insects and certain pathogens. Indeed, Lecanicillium spp. have shown inhibitory effects on a variety of plant pathogens (rusts, mildews, root nematode eggs) while coexisting within the plant or on its surfaces. Harnessing these multifaceted interactions – insect pathogenicity, mycoparasitism, and induced plant resistance – makes V. lecanii a promising agent for holistic crop protection strategies.
Taxonomy, Genetic Tools and Strain Improvement#
Verticillium lecanii has undergone significant taxonomic revision with the advent of DNA-based classification. Originally placed in the genus Verticillium (which largely comprised plant-pathogenic fungi), it became clear in the early 2000s that the entomopathogenic members were genetically distinct. In 2001, Zare and Gams established the genus Lecanicillium to accommodate V. lecanii and related species, based on rDNA ITS sequences and morphological differences from true Verticillium species. Thus, the current accepted name is Lecanicillium lecanii (Zare & W. Gams 2001), and it is recognized as the anamorph (asexual form) of a Cordyceps-like fungus (teleomorph not observed, but DNA places it in the Cordycipitaceae alongside genera like Beauveria and Metarhizium). The genus Lecanicillium has since expanded to dozens of species: many isolates once lumped as “V. lecanii” were split into new species like L. muscarium, L. longisporum, L. nodulosum, etc., each with particular host ranges or traits. This reclassification has practical importance, as commercial strains are now correctly identified (for example, the whitefly-control strain “Ve6” is actually L. muscarium). Ongoing taxonomic research using multigene phylogeny continues to refine the relationships in this group, occasionally placing some Lecanicillium species into other genera like Akanthomyces when warranted. However, Lecanicillium remains a coherent genus for the core entomopathogenic species that share similar conidial structures and life histories.
From a genetics and biotechnology perspective, V. lecanii has become more accessible to laboratory manipulation. Researchers have developed genetic transformation systems for Lecanicillium species to facilitate strain improvement and functional gene studies. One method involves creating nitrate-nonutilizing (nit) mutants and then complementing them with an Aspergillus nitrate reductase gene, allowing foreign DNA integration – this was the basis of a successful transformation of V. lecanii reported in 2011. Stable transformants expressing marker genes (e.g. Green Fluorescent Protein, GFP) have been obtained, enabling tracking of the fungus in host tissues and gene knockout experiments. These genetic tools allow scientists to identify virulence genes (such as those for enzymes and toxins) and enhance desired traits. For example, overexpressing a chitinase gene or a hydrophobin gene (for better spore dispersal) in V. lecanii could yield a strain with faster kill or improved epiphytic survival, respectively. Strain improvement efforts also include classical mutation and selection approaches. Exposure of V. lecanii cultures to chemical mutagens or UV light has yielded variants with higher virulence or wider temperature tolerance. In one study, researchers obtained a hypervirulent strain by screening for faster growth on insect cuticle media. Additionally, protoplast fusion between different Lecanicillium strains has been explored to combine beneficial traits (though interspecific hybrids may be unstable). Modern approaches like genome sequencing are now accelerating strain improvement. The genome of L. lecanii has been sequenced (approximately 32–33 Mb with ~9,500 genes), revealing genes for numerous cuticle-degrading enzymes, secondary metabolite clusters, and stress tolerance mechanisms. By mining these genomic data, researchers can identify targets for genetic enhancement – for instance, upregulating certain toxin biosynthesis pathways to increase insecticidal activity.In parallel, efforts are underway to improve formulation and field performance via strain selection. Traits such as UV resistance, thermotolerance, and shelf stability can vary among V. lecanii isolates. Screening of hundreds of isolates from different climatic regions has identified some that form melanized (pigmented) spores or thicker cell walls, which survive sunlight and heat better. These robust strains are candidates for use in tropical open-field conditions. Biotech companies and agricultural research institutions continue to refine V. lecanii strains through both conventional breeding (screening naturally occurring diversity) and bioengineering, striving for an “ideal” biocontrol strain that combines high virulence, environmental tolerance, and safety.
Challenges and Future Potential#
Despite its many advantages, Verticillium lecanii faces several challenges in broader agricultural use. One major limitation is its dependence on high ambient humidity for effective infection – in dry conditions, the fungus struggles as conidia desiccate or fail to germinate. This means field applications in arid or drought-prone regions have inconsistent results unless irrigation or timing is carefully managed. Additionally, V. lecanii is relatively sensitive to UV light and high temperatures, as noted, which can shorten the persistence of sprayed spores on crop foliage. Another challenge is the relatively slow kill rate compared to chemical insecticides; it may take 5–7 days or more for infected pests to die and cease feeding, during which plants can sustain damage. This slower action requires growers to use V. lecaniipreventatively or in the early stages of pest outbreaks, rather than as a quick knockdown measure. There are also production and formulation challenges: maintaining spore viability in storage (biopesticide products typically have a shelf-life of 6–12 months under cool conditions) and ensuring the spores reach the target pest on the plant. Formulation research is addressing these issues by developing oil-based formulations that stick to leaves and microencapsulated spores that are more shelf-stable.
On the regulatory and adoption front, biopesticides like Lecanicillium face hurdles in registration and grower acceptance. Each strain must be evaluated for safety (to non-target species and consumers) and consistency. Fortunately, V. lecanii has a good safety profile, with no toxicity to mammals, bees, or plants reported in the literature. Some strains have even been tested against beneficial insects and found largely compatible (though heavy spore doses can infect some predatory insects, so timing and selective application are important). The cost of production of V. lecanii biocontrol products can be higher than chemicals, but this is often offset by the benefits of residue-free produce and sustainability. As environmental regulations tighten against harmful pesticides, there is growing future potential for fungi like V. lecanii. Its ability to control multiple pest types (aphids, whiteflies, thrips, etc.) and even plant pathogens positions it as a versatile tool. Future improvements may involve formulating V. lecanii with other biocontrol agents – for example, combining it with compatible predatory insects or parasitoids to achieve immediate and long-term pest suppression (a strategy known as biocontrol augmentation and conservation).
Emerging research avenues include genetically enhancing V. lecanii for stress tolerance, as mentioned, and exploring its metabolites as bio-insecticides. The discovery that fungal proteins can act as plant defense elicitors (see Spotlight study below) opens the door to using V. lecanii not just as a direct insect pathogen but as a sort of biochemical immune boosterfor crops. Another promising direction is using Lecanicillium in seed coatings or soil inoculants so that crops are systemically protected throughout their growth – the fungus could establish in the root zone or within the plant and shield it from both soil pests and foliar pests via induced resistance. In terms of scope, Lecanicillium spp. are being investigated for controlling not only greenhouse pests but also field infestations of pests like coffee green scale, banana aphids, and even certain mosquito species (as larvicides). The adaptability of this fungus could be leveraged to manage pest populations in various ecosystems, provided we address the environmental requirements.In summary, Verticillium lecanii (and its Lecanicillium relatives) has proven to be an effective and eco-friendly biocontrol agent, though optimal use requires understanding its biology. Continued research and development – improving strains, formulations, and deployment strategies – are likely to expand its applicability. With climate change and the push for sustainable agriculture, entomopathogenic fungi like V. lecanii will play an increasingly important role in integrated pest management programs.
Spotlight on Research: Verticillium lecanii Elicitor for Pest Control#
Brief Overview#
In 2021, a team of researchers conducted a notable study to explore Verticillium lecanii from a novel angle – not just as a pathogen of insects, but as a source of biochemical elicitors to enhance plant defense. The study, published in the journal Agriculture, investigated a partially purified protein derived from Lecanicillium lecanii and its effects on cotton plants and whitefly pests. Led by Yusuf A. Abdulle and Talha Nazir (at the Chinese Academy of Agricultural Sciences in Beijing and Ghazi University in Pakistan), the project isolated a protein from L. lecanii culture filtrates and applied it to cotton (Gossypium hirsutum) leaves. The goal was to see if this fungal protein could induce the plant’s own defense mechanisms against the cotton whitefly (Bemisia tabaci), a devastating sap-sucking pest. In essence, the researchers were harnessing a V. lecanii-derived molecule as a plant immune booster, rather than using the live fungus to infect insects directly.
The experimental approach involved treating cotton plants with the Lecanicillium protein at different concentrations and then challenging the plants with whiteflies. They measured outcomes like whitefly survival and reproduction, as well as the expression of key plant defense genes. The study was a collaborative effort spanning institutions in China, Pakistan, and Saudi Arabia, reflecting a multidisciplinary interest in sustainable pest management. By the end of the experiments, the team had gathered evidence that the V. lecanii-derived protein significantly impacted the pest and the plant’s biochemistry. Below we highlight the key insights and implications of this research, which showcases an innovative use of an entomopathogen in crop protection.
Key Insights#
The results of the 2021 study were striking. Cotton plants treated with the L. lecanii protein showed a marked reduction in whitefly fecundity (egg-laying and population growth) compared to untreated controls. Whiteflies exposed to protein-treated plants had lower reproduction rates, indicating a sub-lethal effect of the elicitor on pest fitness. Importantly, the treated plants themselves underwent significant changes: the researchers found that several plant defense-related genes were strongly upregulated. In particular, genes associated with the salicylic acid (SA) signaling pathway (a pathway often activated in response to sap-sucking pests and pathogens) were more strongly expressed in the protein-treated cotton leaves. This suggests that the V. lecanii protein acted as an elicitor, priming the plant’s immune system. The study noted that SA-pathway genes were induced more than jasmonic acid (JA) pathway genes, aligning with the notion that whitefly resistance in plants is typically SA-mediated.
Another key insight was the specificity and potency of the fungal protein’s effect. While many biocontrol strategies focus on killing pests directly, this approach demonstrated that even without causing immediate pest mortality, a fungal metabolite can disrupt pest reproduction and behavior via the plant. The partially purified protein did not harm the cotton plants; on the contrary, it seemed to enhance their defensive state. The authors likened their findings to previous work with Beauveria bassiana elicitors (e.g. a protein named PeBb1 from B. bassiana that has anti-insect effects), reinforcing that entomopathogenic fungi can yield bioactive compounds with pest control potential beyond infection. In summary, the study provided proof-of-concept that V. lecanii can indirectly suppress pests by turning the plant into a less hospitable host.
Why This Matters#
This research is significant because it expands the utility of Verticillium/Lecanicillium in pest management. Traditionally, using entomopathogenic fungi means applying live spores and relying on infection, which, as discussed, can be slow and environment-dependent. The idea of extracting an elicitor molecule from the fungus to induce plant resistance offers a complementary strategy. For farmers, this could translate to a novel type of bio-based “plant strengthener” that reduces pest damage without needing the fungus to actively infect each insect. It also potentially circumvents some limitations of live fungal applications – for example, an elicitor protein could be formulated in a spray that remains effective even in suboptimal humidity, since it doesn’t require fungal growth on the leaf. Moreover, such an approach might have a broader spectrum: induced plant defenses can affect not only the target pest (whiteflies in this case) but also other pests and even microbial pathogens to some extent.From a scientific perspective, the study underscores the multi-faceted roles of entomopathogenic fungi. V. lecanii is not only a killer of insects but also a source of chemical cues that can modulate plant-insect interactions. This bridges the fields of entomology and plant pathology in a novel way. It also suggests a sustainable strategy to manage pests by boosting a crop’s innate immunity, potentially reducing the need for chemical pesticides. If the protein identified in this study (and others like it) can be fully purified and characterized, it might lead to a new class of biopesticide products (sometimes called “biotic elicitors” or “induced resistance agents”). Such products could be used in rotation with traditional biocontrol to achieve both immediate and long-lasting pest suppression.
Summary Table: Spotlight Study#
| Category | Details |
| Lead Researchers | Yusuf A. Abdulle; Talha Nazir (co-first authors); Dewen Qiu (PI) et al. |
| Affiliations | State Key Lab for Biology of Plant Diseases and Insect Pests, CAAS (Beijing, China); Ghazi University (Pakistan); Taif University (Saudi Arabia); others |
| Research Focus | Utilizing a Verticillium lecanii-derived protein to induce plant defenses against whitefly pests (an “elicitor” approach to pest management) |
| Key Breakthroughs | First demonstration that a Lecanicillium protein can trigger salicylic acid-based defense in cotton, significantly reducing whitefly reproduction without direct fungal infection. Validated the concept of fungal secondary metabolites as plant protectants. |
| Collaborative Efforts | International collaboration between institutions in China (CAAS, ICR), Pakistan (universities in Dera Ghazi Khan, Multan, Sargodha), and Saudi Arabia, integrating expertise in entomopathology and molecular plant defense. |
| Published Work | Agriculture (MDPI) journal, Volume 11, Issue 8, article 778 (2021), titled “Sub-Lethal Effects of Lecanicillium lecanii-Derived Protein and Its Potential Implication in Cotton Defense against Bemisia tabaci”. |
| Perspective | Introduces a novel biocontrol paradigm: using entomopathogen-derived elicitors to strengthen crop immunity. Seen as a complementary strategy to direct biopesticide use, potentially leading to new bioproducts that are effective even when environmental conditions do not favor fungal infection. The study’s approach aligns with sustainable agriculture and integrated pest management goals. |
| Publication Date | 15 August 2021 (online publication) |
| Location | Laboratory experiments conducted in Beijing, China (CAAS facilities), with contributions and field context from Pakistani cotton-growing regions (South Punjab) |
| Key Findings | L. lecanii protein treatment caused a significant decrease in whitefly fecundity and triggered upregulation of plant defense genes (especially SA-pathway genes) in cotton. The fungal elicitor did not harm the plant, indicating a viable tool for pest resistance. Authors suggest further purification and characterization of the protein for development as a biocontrol agent. |
Conclusion#
Verticillium lecanii has emerged as a versatile agent in the biological control of agricultural pests, celebrated for its ability to naturally infect and kill a variety of insect pests while being safe to non-target organisms. This “white muscardine” fungus offers a sustainable alternative to chemical pesticides, aligning with eco-friendly integrated pest management practices. In this article, we reviewed the microbe’s biology – from its infection mechanisms (using enzymes like chitinases to penetrate insect cuticles) to its mass production and application in fields and greenhouses. We also explored its secondary benefits, such as endophytic colonization of plants and antagonism against plant pathogens, which broaden its role beyond insect control. Advances in taxonomy and genetics have clarified the identity of V. lecanii (now Lecanicillium lecanii) and equipped researchers with tools to improve its efficacy (through strain enhancement and genetic modification). While challenges like humidity requirements and slower action persist, ongoing innovations in formulation and the discovery of fungal elicitors are addressing these limitations. The spotlight study exemplified an innovative use of V. lecanii – harnessing its bioactive compounds to boost plant defenses, an approach that could complement traditional biopesticide usage.
In conclusion, Verticillium lecanii represents both the rich past and the promising future of microbial pest control. From early greenhouse trials in the twentieth century to cutting-edge molecular research today, this fungus has continually proven its worth as a biocontrol agent. Its ability to adapt (with human guidance) to new roles – whether as a direct insect pathogen, a plant endophyte, or a source of pest-resistance factors – underscores the dynamic potential of entomopathogenic fungi in agriculture. As we refine application strategies and overcome deployment challenges, V. lecanii and its relatives are poised to play an increasingly significant role in securing crops against pests in an environmentally sustainable way.
References#
- Liu, T. et al. (2025). Two novel entomopathogenic fungal species of Lecanicillium isolated from soil in China. BMC Microbiology 25(1): 49. (Introduction provides history, taxonomy and pest control uses of Lecanicillium)
- Zhu, Y. et al. (2008). Isolation and characterization of a chitinase gene from entomopathogenic fungus Verticillium lecanii. Brazilian Journal of Microbiology 39(2): 314–320. (Demonstrates cloning of Vlchit1 chitinase and discusses role of chitinases in fungal virulence)